
Remember me

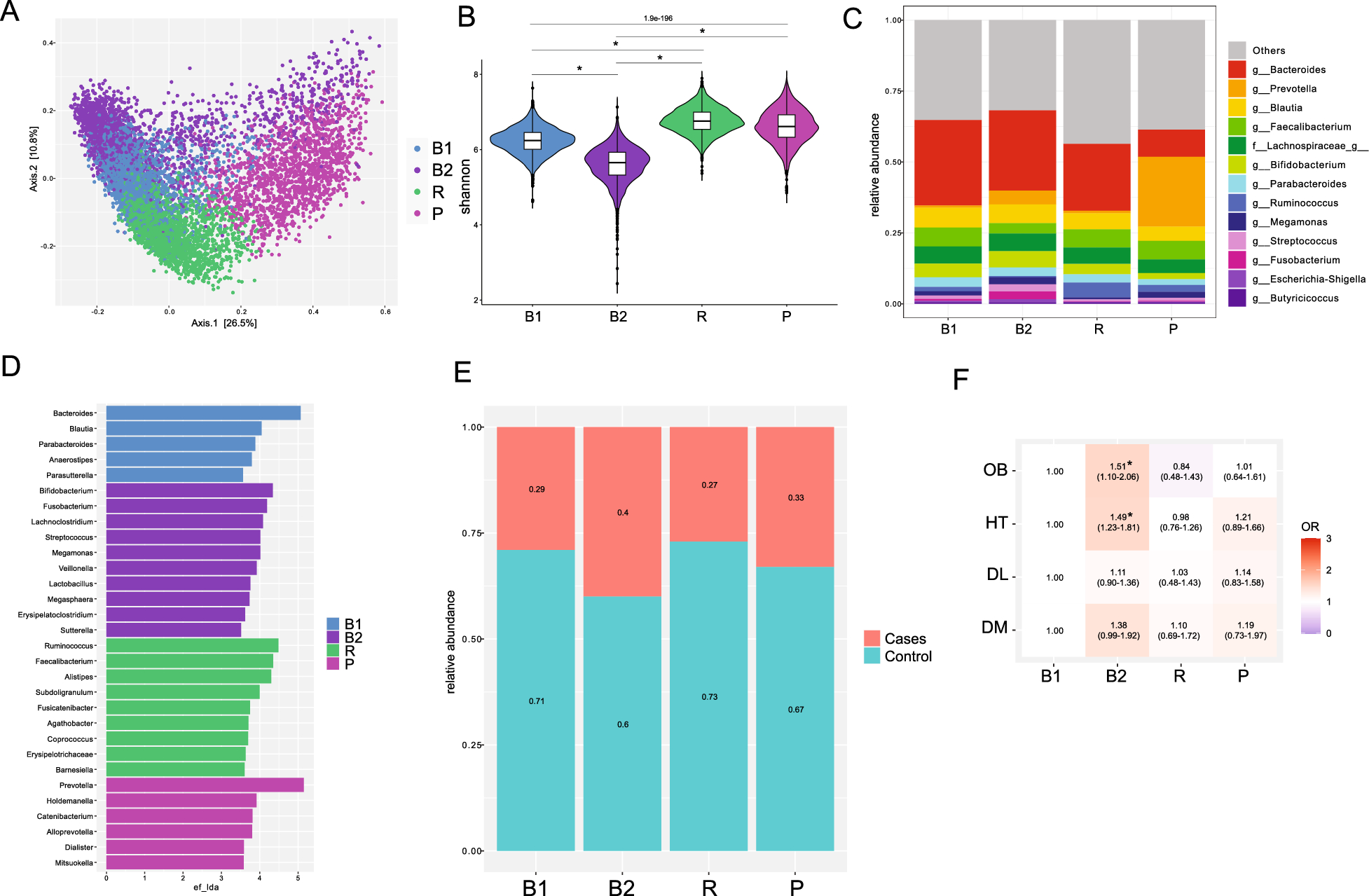
In this study, female Wild-Type (WT) mice and MPO−/− mice were used as research objects to establish a simulated plateau hypoxia model and an inflammation model induced by S. Typhimurium. Mice were initially divided into eight groups: control group (CON), S. Typhimurium infection group (S), hypobaric hypoxia group (H), hypobaric hypoxia plus S. Typhimurium infection group (HS), MPO−/− group (M), S. Typhimurium infection MPO−/− group (MS), hypobaric hypoxia MPO−/− group (HM), and hypobaric hypoxia plus S. Typhimurium infection MPO−/− group (HMS). The body weight changes of mice in each group were monitored daily and it was found that hypoxic exposure and S. Typhimurium infection induced body weight loss. There was no significant difference in the baseline body weight between the WT and MPO−/− groups. Compared with the gradual increase in body weight in the CON and M groups, a pronounced decrease was observed in the H and HM groups (Fig. 1A, B). The reduction in body weight of mice was significantly different from their respective baseline levels in the H and HM groups (Fig. 1A, B). As expected, mice infected with S. Typhimurium exhibited significant body weight loss which was pronouncedly different from their respective baseline levels (Fig. 1A, B). At the same time in the experiment (day 1, day 2, day 3), there were significant differences in body weight between H and CON, HM and M, and HS and S, while there were significant differences in body weight between HMS and MS on day 2 and day 3. Moreover, mice infected with S. Typhimurium (S, HS, and MS groups) exhibited high mortality (100%) on day 5 of infection. In contrast, the MPO−/− mice infected with S. Typhimurium and exposed to hypoxia (HMS group) exhibited more significant early mortality, with 100% mortality reached at 4 days after infection (Fig. 1C). By survival analysis, P = 0.0068 indicates that the four groups of mice infected with S. Typhimurium (S, HS, MS, and HMS groups) are different overall, that is, there are differences between at least two groups. Pairwise comparison was performed among the four groups, between which HMS group and S group (P = 0.0008 < 0.0083), indicating a significant difference, but there is no significant difference between S and MS (P = 0.0382 > 0.0083) and between HS and HMS (P = 0.0785 > 0.0083).
Fig. 1
Hypoxia increases mortality in Salmonella-infected mice. (A) The changes in the body weight in WT mice. (B) The changes in the body weight in MPO−/− mice. Data were analyzed using repeated-measures ANOVA for body weight (n = 8/group; # p < 0.05 versus respectively with their baseline weight; & p < 0.05 versus CON group at the same time; *. p < 0.05 versus S group at the same time; + p < 0.05 versus M group at the same time; ^. p < 0.05 versus MS group at the same time). (C) Mice were continuously monitored for their survival (n = 8 /group). (D) neutrophilic granulocytes, (E) percentage of neutrophils, (F) red blood cell, and (G) hemoglobin were measured at 72 h. (H) Gene expression of VEGF was assessed via qPCR in colon. Values are expressed as mean ± SEM (n = 8 /group) and analyzed via one-way ANOVA with Tukey’s multiple comparisons test (*p < 0.05; **p < 0.01; ***p < 0.001)
The number and proportion of neutrophils in peripheral blood increased during hypoxia (Fig. 1D, E), increased significantly after infection (Fig. 1D, E), and were significantly higher in the MPO−/−-infected mice than in the WT-infected mice (Fig. 1D, E). These results suggest that neutrophil counts and percentages are increased in hypoxia and bacterial diseases.
Hypoxic stress induces erythrocytosis [35]. The RBC and Hb levels in the blood were monitored to verify the hypoxia model (Fig. 1F, G). Hypoxic conditions were determined by induction of vascular endothelial growth factor (VEGF) in the colon (Fig. 1H). The results showed that the red blood cell (RBC), hemoglobin (Hb), and VEGF levels increased after hypoxic exposure (Fig. 1F, G, H), which demonstrated that the hypoxic model was successfully established (Fig. 1F, G, H).
Hypoxia exacerbates bacterial translocation in mice infected with S. TyphimuriumWe hypothesized that high mortality is associated with sepsis. To determine whether hypoxia enhances bacterial migration and whether the increased susceptibility to Salmonella infection in the MPO-deficient mice is related to increased bacterial replication, we investigated bacterial translocation to the spleen and liver at 72 h after infection. No colonies were formed in the simple hypoxia group on LB plates (data not shown). The number of bacterial colonies in the spleen (Fig. 2A) and liver (Fig. 2B) of the hypoxic infection groups was significantly higher than that in the non-hypoxic infection groups (Fig. 2A, B). Moreover, the increase in the MPO−/−-infected mice was more pronounced than that in the WT-infected mice (Fig. 2A, B).
Fig. 2
Hypoxia exacerbates bacterial translocation in S. Typhimurium-infected mice. Bacterial translocation to the spleen (A) and liver (B) was determined 72 h after infection via one-way ANOVA with Tukey’s multiple comparison test. The CFU number per gram of tissue was displayed (n = 8–10/group; *p < 0.05; **p < 0.01; ***p < 0.001). (C) Immunofluorescence microscopy revealed the presence of Salmonella (in green) in spleen sections (×200). The spleen (D) and liver (E) were stained with hematoxylin and eosin [(D) ×200; (E) ×400]. The blue arrow represents lymphocyte hyperplasia. The black arrow means red marrow congestion. The yellow arrow indicates that the structure of the liver plate is unclear. The green arrow represents focal abscess. (F–I) Relative mRNA expression of TNF-α (F, H) and IL-1β (G, I) in the spleen and liver tissues was detected by qPCR. (J-M) Levels of TNF-α (J, L) and IL-1β (K, M) were detected by ELISA. Data are presented as mean ± SEM (n = 8 /group) and analyzed via one-way ANOVA with Tukey’s multiple comparisons test (*p < 0.05; **p < 0.01; ***p < 0.001)
This was further supported by immunofluorescence microscopy analysis of the spleen tissue (Fig. 2C), which suggested that hypoxia increased bacterial translocation and MPO-deficient mice had enhanced bacterial dissemination in the spleen tissue. An increase in tissue bacterial load is accompanied by worsening inflammation and tissue damage. Histological analysis of the spleen tissue showed elevated lymphocyte levels in the HM group, indicating cellular destruction (Fig. 2D). The S and MS groups showed extensive spleen abnormalities, as evidenced by red pulp congestion, neutrophil infiltration, and abscess formation (Fig. 2D). The splenic tissue of infected mice under hypoxic condition, HS and HMS groups showed obvious hyperemia red pulp congestion, multiple small abscess foci, and splenic architecture disruption. The MPO−/−-infected group was more severely affected than the WT-infected group (Fig. 2D).
Concurrent with the above analyses, liver histomorphology was observed. Histological analysis showed that the hepatocytes in the HM group were disarranged and the hepatic plate was unclear (Fig. 2E). Extensive necrosis and mixed inflammatory cell infiltration were observed in both WT-infected and MPO−/−-infected liver sections. The liver lesions in the WT-infected mice were characterized by neutrophil infiltration and small abscess formation. MPO−/−-infected mice showed extensive neutrophilic infiltration with histiocytic and lobulated neutrophils, and multi-focal small abscesses. Liver lesions in WT and MPO−/− mice with hypoxic infection also showed extensive neutrophil infiltration and multi-focal small abscess formation (Fig. 2E). These results suggest that there is significant inflammation and tissue damage in the spleen and liver of both WT mice and MPO−/− mice after Salmonella infection because of the higher bacterial populations present in these tissues.
These findings were consistent with the more severe inflammatory changes in the spleen and liver. Salmonella-infected WT and MPO−/− mice showed increased mRNA expression of proinflammatory mediators, including TNFα (Fig. 2F, H) and Il-1β (Fig. 2G, I). Hypoxia significantly increased the TNFα and Il-1β mRNA levels in the MPO−/− mice compared to that in the WT mice (Fig. 2F, G, H, I). In addition, the expression of these two genes was significantly increased in the Salmonella-infected MPO−/− mice compared to that in the WT-infected mice (Fig. 2F, G, H, I). Furthermore, the concentration of TNF-α (Fig. 2J, L) and IL-β (Fig. 2K, M) in the spleen and liver homogenates analyzed by ELISA were consistent with the expression of mRNA. These results suggest that hypoxia leads to increased production of proinflammatory cytokines, which are overexpressed due to MPO deficiency in hypoxia or infection conditions and may be attributed to the observed aggravated tissue damage. These findings indicate that oxygen and MPO play a vital role in host resistance and survival during Salmonella infection.
Hypoxia enhances macrophage and neutrophil recruitment in S. Typhimurium-infected miceTo further study the effects of hypoxia and MPO on the innate immunity against Salmonella infection, we analyzed the spleen tissue’s frequency of neutrophils and macrophages. FACS analysis revealed that the spleen cells had a high number of macrophages that expressed CD11b+ F4/80+ in the MPO−/− mice compared to that in the WT mice exposed to hypoxic conditions (Fig. 3A, C). Compared with WT-infected mice, the number of macrophages expressing CD11b+ F4/80+ was increased in MPO−/−-infected mice, and the number of macrophages expressed in MPO−/−-infected mice was significantly increased in comparison to the number in WT-infected mice under hypoxic conditions. (Fig. 3A, C). The frequency of CD11b+ Ly6G+ neutrophils in the spleens of the mice was consistent with the number of macrophages (Fig. 3B, D). These data suggest that hypoxia and MPO deficiency promote the migration of neutrophils and macrophages to the infection site. Next, we determined the expression of three chemoattractant molecules ((KC (CXCL1), MCP1 (CCL2), and MIP2 (CXCL2)) that regulate neutrophil and macrophage recruitment in spleen tissue. RT-qPCR results showed the hypoxia-induced upregulation of KC, MCP1, and MIP2 gene expression, and their expression in the MPO−/− mice was significantly higher than that in the WT mice (Fig. 3E, F, G). Salmonella infection augmented the expression of these genes (Fig. 3E, F, G). The increase in chemokine expression was most apparent in MPO-deficient mice compared to the WT mice after infection (Fig. 3E, F, G). These results suggest that hypoxia and MPO deficiency may promote the recruitment of phagocytes to the site of infection by regulating the expression of chemokines, and that excessive aggregation of granulocytes in the spleen may enhance the inflammatory response during Salmonella infection.
Fig. 3
Hypoxia enhances neutrophil and macrophage recruitment in mice during Salmonella infection. Single-cell suspensions of spleens were prepared and stained with anti-CD11b and anti-F4/80 antibodies for macrophages (A) and with anti-CD11b and anti-Ly6G for neutrophils (B). The percentages of CD11b+F4/80+ macrophages (C) and CD11b+ Ly6G+ neutrophils (D) are shown as mean ± SEM (n = 6/group). (E-G) Gene expression of chemoattractant (KC, MCP1, and MIP2) in spleen was analyzed via qPCR (n = 8/group). Significance was analyzed by one-way ANOVA with Tukey’s multiple comparison test (*p < 0.05, **p < 0.01, ***p < 0.001)
Hypoxia augments Salmonella-induced mucosal injury in miceBacterial translocation is generally the result of impaired gut integrity. Our results suggest that hypoxia induces more severe liver and spleen infections in MPO-deficient mice during Salmonella infection. We investigated whether hypoxia and MPO levels affect innate immunity and inflammation in the intestinal mucosa. Under a light microscope, colon morphology was examined, and the results showed that the colonic mucosa had an intact epithelium and a smooth appearance in the CON and M groups (Fig. 4A). Compared with the above two groups, the recesses of the colon in the H and HM groups were shallower and flatter. Inflammatory cell infiltration was observed in the S and MS groups (Fig. 4A). The HS group showed obvious inflammatory cell infiltration and necrosis; however, this was more severe in the HMS group. Inflammatory cell infiltration with suppuration was observed in the colon of the HMS group (Fig. 4A). Immunofluorescence microscopy analysis of infected colons revealed an increased bacterial load in hypoxia-infected (Fig. 4B) and MPO-deficient mice compared to that in the WT mice (Fig. 4B). In addition, consistent with intestinal injury, hypoxia increased the levels of TNF-α (Fig. 4C) and IL-β (Fig. 4D) in the MPO−/− mice compared to those in the WT mice. The mRNA expression levels of TNF-α (Fig. 4C) and L-1β (Fig. 4D) were significantly increased in the colon of MPO−/−-infected mice compared to that in the WT-infected mice (Fig. 4C, D). ELISA analysis of cytokine concentrations in the colon tissue was consistent with those of mRNA expression (Fig. 4E, F).
Fig. 4
Hypoxia augments Salmonella-induced mucosal injury and colitis in mice. (A) Hematoxylin and eosin staining of the colon (×100, ×400). (B) Immunofluorescence microscopy data show the distribution of Salmonella (in green) in colon sections (×200). Levels of TNF-α (C) and IL-1β (D) were determined by quantitative PCR. Cytokine levels of TNF-α (E) and IL-1β (F) were detected by ELISA. Colon levels of KC (G), MCP1 (H), and MIP2 (I) were analyzed by qPCR. Data represent mean ± SEM (n = 8 /group) and one-way ANOVA followed by Tukey’s multiple comparisons test (*p < 0.05; **p < 0.01; ***p < 0.001)
Next, we measured chemokine mRNA levels in the colon tissue. The results showed that hypoxia boosted the expression of KC, MCP1, and MIP2 in the colons of mice, especially in the MPO−/− mice (Fig. 4G, H, I). Salmonella infection promoted chemokine expression in colon tissue (Fig. 4G, H, I), and the upregulated chemokines were significantly different in both WT-infected and MPO−/−-infected mice. Compared with WT-infected mice, the expression of chemokines (KC, MCP1, and MIP2) in MPO−/−-infected mice was more significant (Fig. 4G, H, I). These results suggest that Salmonella-induced inflammation can be demonstrated in both WT mice and MPO−/− mice which can be exacerbated by hypoxia. Moreover, MPO−/− mice present aggravated Salmonella-induced inflammation.
Hypoxia aggravates colonic oxidative stress injury in MPO-deficient miceOxidative stress is associated with the progression of various inflammatory diseases of the colon, and the lack of antioxidant defenses in the intestinal mucosa is an essential factor leading to the impairment of the intestinal mucosal barrier [36, 37]. MPO activity is an inflammatory biomarker. Therefore, the activities of malondialdehyde (MDA), catalase (CAT), glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), and MPO were measured in the colon tissue. The results showed that the activity of MDA was significantly increased and the activities of CAT, GSH-Px, and SOD were remarkably decreased under hypoxic conditions (Fig. 5A, B, C, D). The findings for MPO−/− mice differed significantly from those of WT mice under hypoxic conditions (Fig. 5A, B, C, D). The activities of MDA, CAT, GSH-Px, and SOD in colon tissue after Salmonella infection were similar to those under hypoxia (Fig. 5A, B, C, D). Moreover, the MPO−/−-infected mice showed higher MDA activity but lower CAT, GSH, and SOD activities than the WT-infected mice (Fig. 5A, B, C, D). The level of intestinal MPO was increased in the WT mice under hypoxic conditions and increased significantly after infection (Fig. 5E). Next, a ROS assay was performed on frozen sections of colon tissue. The results showed that hypoxia and infection increased the mean fluorescence intensity of ROS (Fig. 5F, G). ROS production in the Salmonella-treated MPO−/− mice was higher than that in the Salmonella-treated WT mice (Fig. 5F, G). The results suggest that hypoxia increases ROS levels, and MPO deficiency aggravates oxidative stress.
Fig. 5
Hypoxia aggravates oxidative stress injury in colon during Salmonella infection Colon activity of MDA (A), GSH-Px (B), SOD (C), CAT (D), and MPO (E) were determined (n = 8 /group). (F-G) Mean fluorescence intensity of ROS was determined (×200) (n = 3/group). (H) Western blot performed and (I-O) quantification (n = 3/group). Data are presented as mean ± SEM and analyzed via one-way ANOVA with Tukey’s multiple comparisons test (*p < 0.05; **p < 0.01; ***p < 0.001)
The protein expression levels of key molecules in the Nrf2/HO-1 pathway were determined. The protein levels of Nrf2 and HO-1 were significantly increased under hypoxia, especially in WT mice (Fig. 5H, I, J). After Salmonella infection in WT mice, the HO-1 protein levels decreased significantly, and the Nrf2 protein was decreased but not significantly (Fig. 5H, I, J). The protein levels of Nrf2 and HO-1 were significantly lower in the Salmonella-infected MPO−/− mice compared to those in the WT-infected mice (Fig. 5H, I, J). In addition, the Salmonella-treated WT and MPO−/− mice presented increased levels of proteins, including iNOS, NLRP3, p-65/T-NF-kB p65, and p-ERK/T-ERK (Fig. 5H, K, L, M, N). Hypoxia significantly increased these levels in the MPO−/− mice compared to those in the WT mice (Fig. 5H, K, L, M, N). Moreover, these protein levels were significantly increased in the Salmonella-infected MPO−/− mice compared to those in the WT-infected mice (Fig. 5H, K, L, M, N). In WT mice, the protein expression level of MPO was significantly increased under hypoxia, and Salmonella-infected mice showed increased protein levels (Fig. 5H, O).
Comments (0)