
Remember me


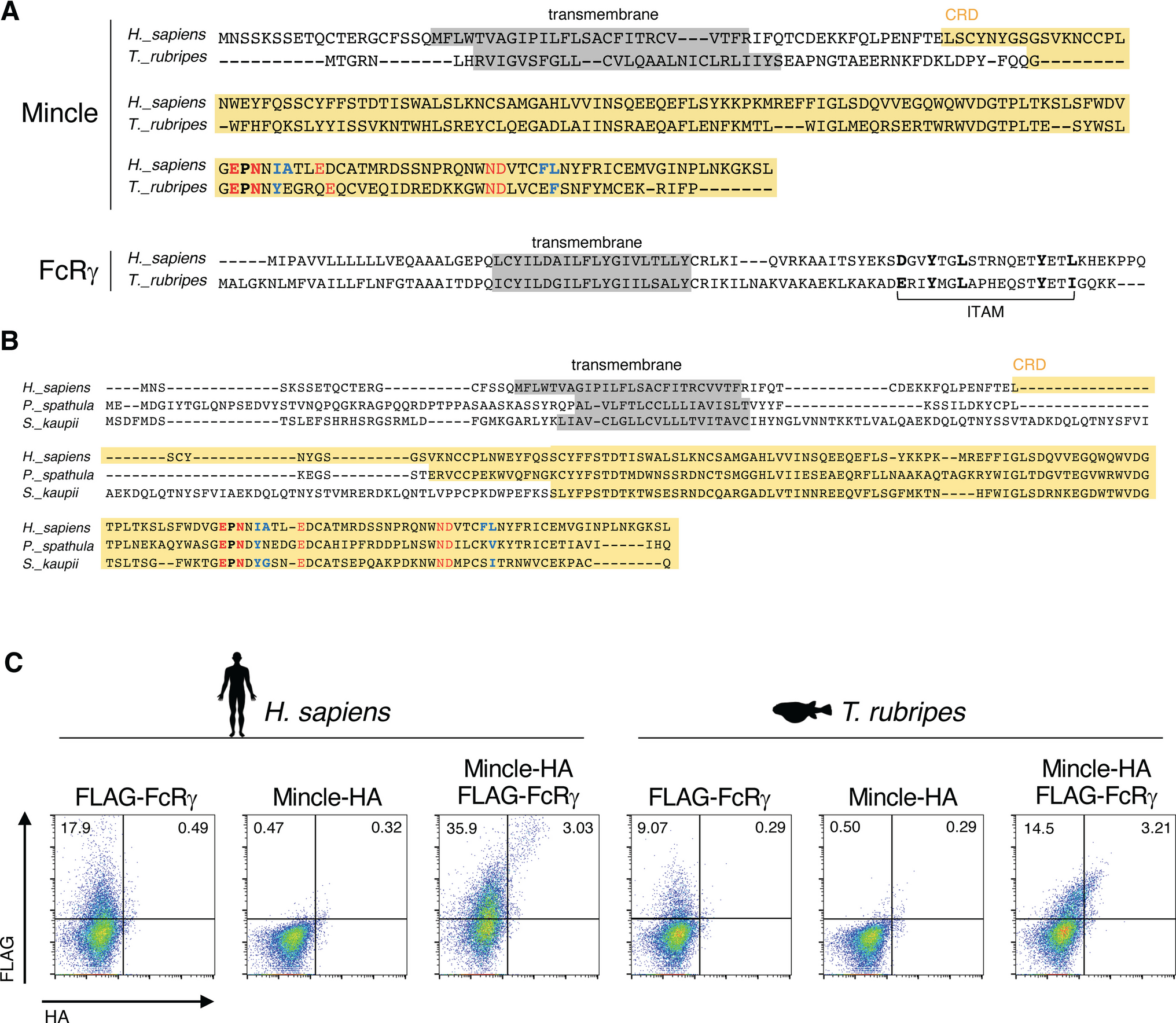
Recombination is an important mechanism in genome evolution, which manifests in a variety of ways. The relative organisation of genes and sequence features can have an effect on the outcome of recombination in a multigene family (Fig. 5).
Examples of different outcomes of recombination have been found in the chicken B locus, which includes the BG genes in the BG region and the chicken MHC in the BF-BL region. For homologous recombination between similar sequences in the same orientation (that is, small direct repeats or homologous genes that are in the same transcriptional orientation) but on different chromosomes, the result can be hybrid genes with resulting expansion or contraction of the gene family (Fig. 5a), as is seen for the multigene family of BG genes (Salomonsen et al. 2014). For such homologous recombination between direct repeats on the same chromosome, the result can be deletion (Fig. 5c), as is seen for the BF1 promotor in the B12 and B19 MHC haplotypes (Shaw et al. 2007) and for the 5′ untranslated regions (5′UTRs) of tissue-expressed BG genes (Salomonsen et al. 2014).
For such homologous recombination on the same chromosome between genes in opposite transcriptional orientation or between inverted repeats, the result can be inversion (Fig. 5d, e), as may be the case for the BLB1 and BLB2 genes found in the chicken MHC (Afrache et al. 2020). Indeed, the fact that several pairs of genes with similar sequences are in opposite transcriptional orientation (BF1 and BF2, TAP1 and TAP2, BLB1 and BLB2, BNK and Blec, Fig. 6) has been interpreted as a way to prevent loss of essential genes from the compact and simple chicken MHC (Afrache et al. 2020; Kaufman 2023), originally described as a “minimal essential MHC” (Kaufman et al. 1995, 1999). Although such inversions have yet to be conclusively demonstrated for genes in the chicken MHC, the orientation of tapasin and TAP genes relative to flanking genes in closely related avian species (Fig. 6) is consistent with such inversion (He et al. 2021), as previously noted (Kaufman 2023).
Fig. 6
The chicken MHC (BF-BL region) with all the published genes compared to the chicken MHC with the BF1 deletion and with same regions in closely related birds, showing that the transcriptional orientation of the tapasin, TAP1, and TAP2 genes can reverse between species, most easily explained by inversion of flanking genes in opposite transcriptional orientation. Solid lines indicate genomic DNA, boxes indicate genes (red, class I system; blue, class II system; green, other immune genes; white, serine/threonine kinase gene BRD2; slanted lines, genes involved in peptide loading) with names above (except for BF1, BF2, BLB1 and BLB2 genes in chickens whose characteristics are well-described, the class I and class II B genes of other species indicated by BF and BLB, respectively), and transcriptional orientations indicated by arrows below (bold red indicates a switch compared to chickens). Double-headed arrows indicate homologous gene pairs in chickens. Figure modified from Kaufman et al. 1999) with data from this paper and from He et al. 2021)
However, despite the fact that several key gene pairs in the chicken MHC are in opposite transcriptional orientation which would be expected to protect against deletion, the lack of BF1 cDNA in two standard haplotypes, B14 and B15, was noted long ago (Kaufman et al. 1995; Wallny et al. 2006; Shaw et al. 2007). The attempts to understand the basis of this lack of expression at the level of genomic DNA by PCR using many primers that were successful for all other haplotypes led to no amplification of bands and by Southern blots led to the identification of much larger bands (Shaw et al. 2007). Two interpretations of the Southern blot data were that there had been changes in the restriction enzyme sites between haplotypes or that there had been an insertion. No changes in the sites were found in the sequences which were then available, which led to the tentative conclusion that an insertion was responsible, but no evidence for the inserted sequence was ever found.
In the present study using amplification of much longer DNA fragments, we find that a large deletion between the promoter of the BF1 gene and the end of the TAP1 gene is responsible for the lack of BF1 expression in the B14 and standard B15 haplotypes and that a direct repeat found in the correct positions in an atypical B15 haplotype is responsible for the deletion. Compared to the direct repeats in the BF1 promoter of the B12 and B19 haplotypes and the 5′UTRs of the tissue BG genes (Shaw et al. 2007; Salomonsen et al. 2014), the direct repeat is longer (32 compared to 10 and 27 nucleotides) and is also more imperfectly repeated (10 compared to 0 and around 3 ambiguities).
The deletion of a presumed “essential gene” in two standard haplotypes (found in perhaps billions of domestic chickens) was unexpected, so how commonly would such a deletion occur? All the BF1 genes examined should be susceptible to this deletion (in contrast to the BF2 genes), due to the presence of exactly the same repeat sequences (except for a few nucleotides in B2), and one of these repeats is located adjacent to the repeat associated with the deletion in the promoter of the B12 and B19 haplotypes (Fig. S1), perhaps indicating that the chromatin might be more open for recombination around promoter sites. However, the promoter deletion apparently happened only once (since the BF1 gene in B19 is derived from the B12 haplotype), and only two of the standard haplotypes (B14 and B15) have deleted the BF1 gene entirely. Moreover, the deletion in the B14 and typical B15 haplotypes appears identical (Fig. 3, S1). Indeed, the 14 single nucleotide polymorphisms (SNPs) in the sequences of B14, standard B15, and unusual B15 haplotypes compared to other haplotypes (Fig. S1) are spatially segregated, with SNPs unique to B14 and SNPs shared between the two B15 haplotypes found in the DM gene and the beginning of the intergenic region, followed by a cluster of SNPs that are shared between all three haplotypes, and finally two SNPs and the big deletion that are shared between B14 and the standard B15 haplotype. Together, the final two SNPs and big deletion might be taken as evidence that there was a single deletion in one haplotype that was transferred to the other haplotype by homologous recombination. However, in the absence of a greater number of informative features, it remains possible that there were independent deletions in the two haplotypes.
Another approach to determine the frequency of similar but independent deletions would be to examine a variety of other MHC haplotypes. An ongoing typing exercise has identified nearly 250 MHC haplotypes in chickens, of which 28 lack an obvious BF1 gene and four appear to lack one BLB gene (Tregaskes et al. 2023). A closer look at these other haplotypes is necessary to determine whether these apparent BF1 deletions have the same basis as the ones described here. Moreover, the potential effects of such “natural gene knock-outs” on resistance to infectious pathogens such as Marek’s disease virus have never been assessed.
Comments (0)