
Remember me

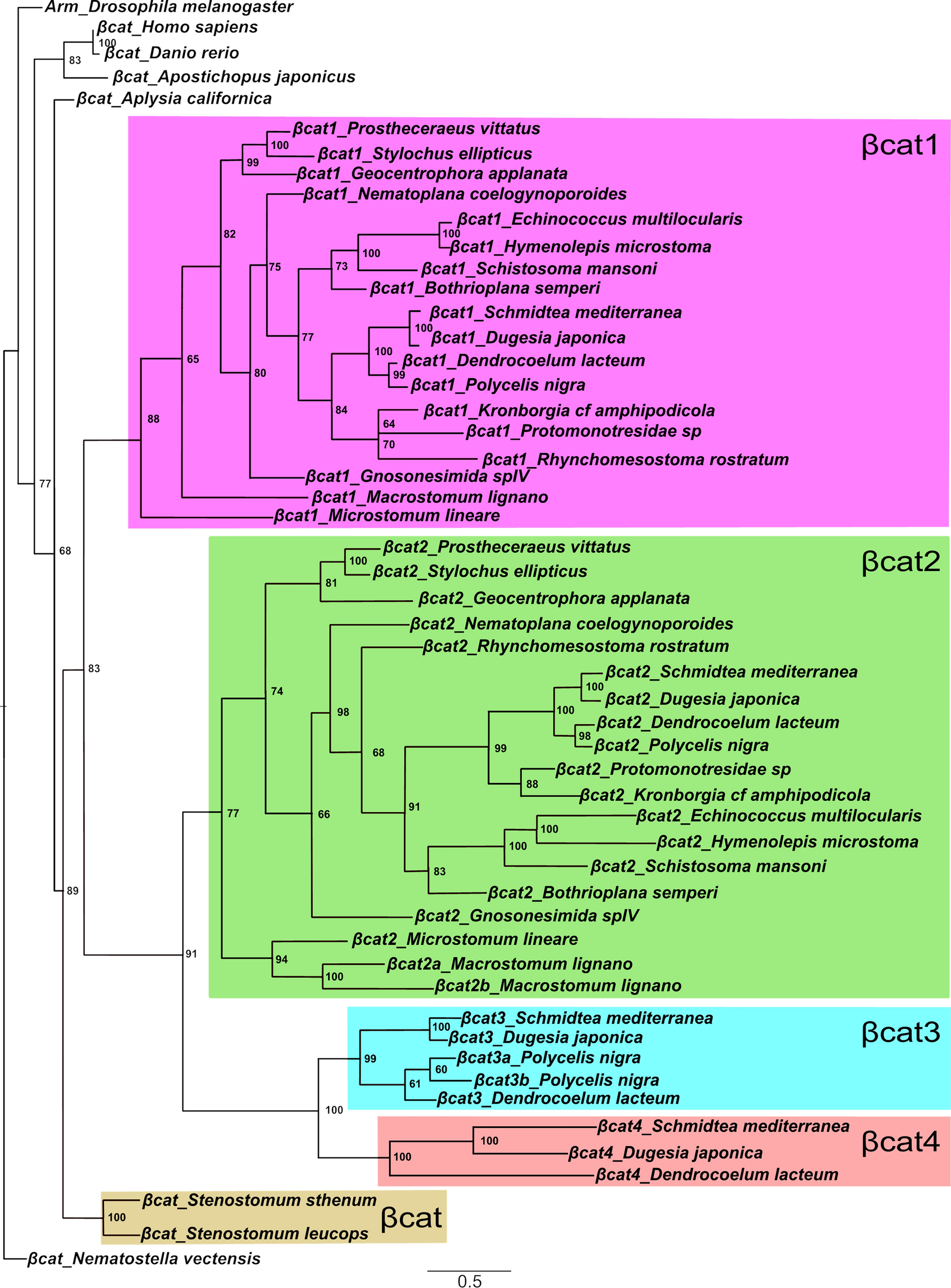

To determine whether inpp5ka and inpp5kb lead to functionally divergent proteins, we first analyzed their protein sequence. Protein sequence alignment of INPP5K (NP_057616.2), Inpp5ka (NP_001082962.2), and Inpp5kb (XP_021335021.1) revealed 45.8% and 43.9% identity between the human orthologue and Inpp5ka and Inpp5kb respectively, while the zebrafish proteins showed 62.2% identity with each other (Fig. 1). Higher conservation was present in the phosphatase domain with 49.7% amino acid sequence identity between the human protein and either zebrafish protein. It is important to note that while the primary isoforms listed above are the most similar to the human gene, both inpp5ka and inpp5kb have additional predicted transcripts that are not found in humans. One transcript has an alternative exon 1 (inpp5ka: XM_009291785.3 and inpp5kb: XM_021479345.1) adding an N-terminal sequence of 63 amino acids for Inpp5ka and 48 amino acids for Inpp5kb. A small 27 base pair alternative exon 9 was also identified in predicted transcripts both inpp5ka (XM_00557623.4) and inpp5kb (XM_005155275.4). All expression and functional analyses performed in this study were based on the sequence of the primary isoforms most similar to the human gene, but the design for the primer and probes did not exclude other predicted isoforms.
Fig. 1
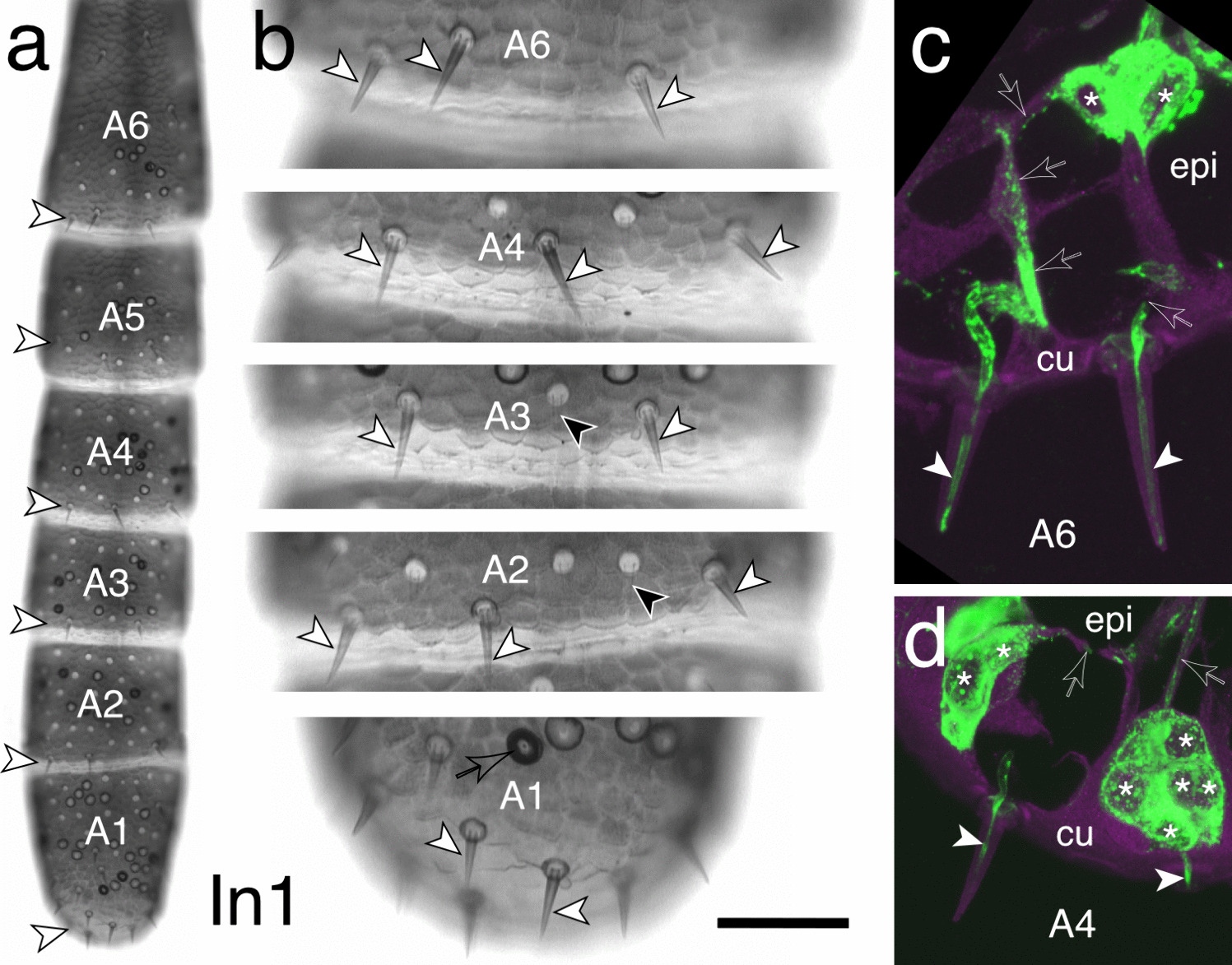
Protein alignment of INPP5K, Inpp5ka, and Inpp5kb highlighting conserved amino acids required for phosphatase activity. The start and end of the catalytic domain in the human protein are marked with large red arrowheads. Amino acids required for phosphatase activity have been denoted with asterisks (*). Small arrowheads indicate residues that are altered by missense variants in humans. Hs, Homo sapiens; Dr, Danio rerio
Divergent expression and localization of INPP5K orthologs in zebrafish larvaAnalysis of inpp5ka and inpp5kb mRNA obtained from whole zebrafish embryos had shown higher expression of inpp5ka (Osborn et al. 2017). We used qPCR to quantify expression patterns throughout the first five days of development. We found that inpp5ka (NM_001089493.1) was consistently expressed much more abundantly than inpp5kb (XM_021479346.1) (Fig. 2A). The developmental expression trend was similar for inpp5ka and inpp5kb. For comparison across the two genes fold changes for the whole embryo were quantified compared to inpp5kb expression at 1 dpf since it showed the lowest levels. Both genes showed relatively low levels at 1 and 2 dpf, but expression increased after 3 dpf and we saw 5.8, 3.2, and 3.6 higher expression of inpp5ka at 3, 4, and 5 dpf, respectively (Fig. 2A, fold change calculated to 1 dpf inpp5kb. inpp5ka: 1 dpf 8.5 ± 0.8, 2 dpf 11.1 ± 1.5, 3 dpf 51.9 ± 0.4, 4 dpf 112.1 ± 3.6, 5 dpf 84.9 ± 5.7; inpp5kb: 2 dpf 2.1 ± 0.1, 3 dpf 9.0 ± 2.3, 4 dpf 35.5 ± 3.9, 5 dpf 23.6 ± 2.5. p > 0.0001 at 3, 4, and 5 dpf).
Fig. 2
inpp5ka and inpp5kb mRNAs differ in expression levels in zebrafish larvae. A Gene expression determined by qPCR. inpp5ka is more highly expressed in whole body lysates through 5 dpf. B Larval tissues were excised from the eye, head, and tail for localized gene expression analysis excluding the area of the trunk around the yolk. C, D Expression for both inpp5ka (C) and inpp5kb (D) is low in the tail and increases in the eyes and brain. By 5 dpf, both are most highly expressed in the eyes. Results from the whole body from (A) are shown as reference. E, F inpp5ka (E) maintains higher expression levels than inpp5kb (F) in the tail/muscle and head at 30 dpf and in adult fish. Both genes are expressed at very high levels in the eyes. Values are averages ± SEM. 2-way ANOVA results for tissue and developmental timepoint (time) are also listed. *p < 0.05, **p < 0.01, ***p < 0.001
Loss of INPP5K in humans affects the muscle, brain, and eyes and knockdown of inpp5ka in zebrafish larvae resulted in morphological abnormalities in the eyes and skeletal muscle (Osborn et al. 2017; Wiessner et al. 2017; Hathazi et al. 2021). We dissected the heads, eyes, and tails of developing larvae for tissue-specific expression analysis (Fig. 2B). Tissue-specific analysis revealed that, while inpp5ka was consistently expressed at higher levels than inpp5kb, both paralogs exhibit the greatest expression in the eyes, intermediate expression in the head, and low levels in the tail. For comparison, we calculated fold changes compared to expression levels of inpp5kb at 1 dpf. inpp5ka expression increased dramatically in the eyes by 5 dpf (Fig. 2C). Both genes showed similar increases in the head by 4 dpf with tail levels remaining consistently low (Fig. 2C, D, fold change relative to 1 dpf inpp5kb in the tail. inpp5ka: 3 dpf head 145.2 ± 19.3, eyes 80.4 ± 31.6; 4 dpf head 271.2 ± 41.5, eyes 528.1 ± 129.8; 5 dpf 218.5 ± 2.7, eyes 653.1 ± 99.6. inpp5kb: 3 dpf head 30.4 ± 3.4, eyes 28.6 ± 4.4; 4 dpf head 217.3 ± 128.8, eyes 157.7 ± 8.4; 5 dpf head 85.8 ± 7.9, eyes 276.9 ± 40.6). Since expression levels in the developmental time course seemed to be dropping after 4 dpf, we asked whether tissue-specific expression differences were still present in juvenile (30 dpf) and adult (1 year). Both inpp5ka and inpp5kb showed very high expression levels in the eyes which were comparable at both ages (Fig. 2E, F). mRNA expression in the head was higher for inpp5ka and increased in adulthood. While expression in the tail at 30 dpf and in dissected muscle at 1 year was low for inpp5ka, but still 7.6 to 11 times higher than inpp5kb (Fig. 2E, F, fold change relative to 30 dpf inpp5kb tail. inpp5ka: 30 dpf tail 7.6 ± 1.4, head 33.9 ± 6.9, eyes 828.9 ± 320.6; 360 dpf tail 11.3 ± 2.1, head 114.5 ± 21.0, eyes 965.9 ± 83.8. inpp5kb: 30 dpf head 16.3 ± 7.1, eyes 1967.1 ± 484.7; 360 dpf tail 0.99 ± 0.22, head 19.6 ± 5.1, eyes 1597.9 ± 615.8). Overall, inpp5ka showed consistently higher levels of expression than inpp5kb throughout the body apart from the eyes where expression of both genes was the highest at juvenile and adult timepoints.
To confirm the expression patterns, we first conducted in situ hybridization on whole-mount larvae at 3 dpf when expression begins to diverge. inpp5ka antisense probes reflected the results of qPCR expression assays. inpp5ka mRNA was most abundant in the head and eyes, with lower expression in the tail (Fig. 3A–C). As expected, inpp5kb antisense targeting revealed lower expression throughout the head and eyes (Fig. 3D, E). However, in contrast with inpp5ka, inpp5kb was abundantly expressed in the pineal gland (Fig. 3F), a neuroendocrine organ which responds to light and plays a role in circadian rhythm (Cahill 1996; Vatine et al. 2011; Livne et al. 2016). These findings suggested that in addition to lower expression, inpp5kb may have also diverged in its expression pattern. To better define the cellular distribution of the two mRNAs we then performed fluorescent RNA scope in situ on tissue cryosections at 5 dpf when expression levels are significantly increased via qPCR. This analysis revealed additional differences. inpp5ka was evenly distributed throughout the brain, in all layers of the retina, and was also present around the nuclei of lens cells (Fig. 4A, B). Expression in the muscle at the same exposure was much lower reflecting qPCR results (Fig. 4C). inpp5kb showed striking differences in distribution with very high expression in the pineal gland (Fig. 4D, Suppl.Fig. 1A) and in the inner nuclear layer of the retina (Fig. 4E, Suppl. Fig. 1B), though expression was still noted in the brain, other retinal layers, and the lens. As seen for inpp5ka, inpp5kb expression in the muscle was very low (Fig. 4F) though nuclei are sparse in this tissue and staining is less concentrated than in the brain and eyes (Suppl.Fig. 1C). Thus, the mRNA expression levels of inpp5kb in the brain and eye appear to be primarily driven by specific expression patterns while inpp5ka has a more even distribution pattern.
Fig. 3
inpp5ka and inpp5kb mRNAs differ in localization at 3 dpf. A–C In situ hybridization in 3 dpf larvae shows that inpp5ka mRNA is highly expressed throughout the head and eyes. Scale bars: 500µm in (A), 100µm in (C). D–F inpp5kb expression is concentrated to the pineal gland. The pineal gland is indicated by the black arrow
Fig. 4
Cellular distribution of inpp5ka and inpp5kb at 5 dpf by RNA scope in situ. A–C inpp5ka shows even distribution in the brain and eye (A, B), also labeling lens cells (B). Muscle expression is sparse and at lower levels (C). D–F inpp5kb brain expression is highest in the pineal gland (arrow in D). Increased expression is also noted in the inner nuclear layer of the retina with lower expression in photoreceptors, retinal ganglion cells, and lens cells (E, also see Suppl. Figure 1B). Sparse expression is present in muscle cells (F, also see Suppl Fig. 1C). G–I Merged images including both probes (inpp5ka in green and inpp5kb in red) and DAPI to counterstain the nuclei. Scale bars: 100 µm for (A), (D), and (G), 50 µm for all other panels
Divergence in phosphatase activity of human and zebrafish orthologs of INPP5KTo evaluate the preservation of the PI phosphatase activity in the zebrafish isoforms, we conducted a malachite phosphatase assay to examine the activity of INPP5K and the two zebrafish Inpp5k isoforms against PIP3 and the preferred substrate PtdIns(4,5)P2 (Fig. 5A). We found that zebrafish Inpp5ka and human INPP5K were both highly active against PtsIns(4,5)P2 as expected. This activity was specific, as illustrated by the lack of phosphatase activity against PIP3. However, compared to Inpp5ka, Inpp5kb was nearly inactive against PtdIns(4,5)P2. Inpp5ka yielded 409 pmol of free phosphate vs 20 pmol for Inpp5kb, indicating that Inpp5ka had a 20-fold higher activity compared to Inpp5kb (Fig. 5B).
Fig. 5
Inpp5ka and Inpp5kb exhibit different phosphatase activity. A Phosphatase activity of human INPP5K, Inpp5ka, and Inpp5kb in the malachite assay. Human INPP5K and Inpp5ka demonstrate high activity for the PI(4,5)P2 substrate. PIP3 did not elicit activity from any isoform. B Inpp5ka is more significantly active against diC8PI(4,5)P2 compared to Inpp5kb. Values are averages ± SEM. ***p < 0.001 following a t-test
The INPP5K protein is primarily composed of a 5-phosphatase domain between amino acids 16–318 and a SKITCH domain between amino acids 321–448. Most mutations identified in humans are missense and have been shown to reduce or ablate phosphatase activity (Osborn et al. 2017; Wiessner et al. 2017). We wondered whether the loss in activity in Inpp5kb could be caused by changes in amino acids identified to be critical for the catalytic activity of INPP5K. Basing this analysis on the available crystal structures of other Type II inositol phosphate 5-phosphatases, INPP5B and SYNJ1 (Trésaugues et al. 2014; Paesmans et al. 2020), we found that all sites were conserved in Inpp5ka and Inpp5kb and there were no major changes that could explain differences in activity (asterisks in Fig. 1). We also assessed whether residues known to be affected by pathogenic variants in patients were conserved in Inpp5kb, and these amino acids were all maintained (small arrowheads in Fig. 1) (D’Amico et al. 2020; Osborn et al. 2017; Hathazi et al. 2021; Wiessner et al. 2017; Yousaf et al. 2017). Thus, possible changes in known residues do not explain the difference in function between Inpp5ka and Inpp5kb.
Comments (0)