
Remember me


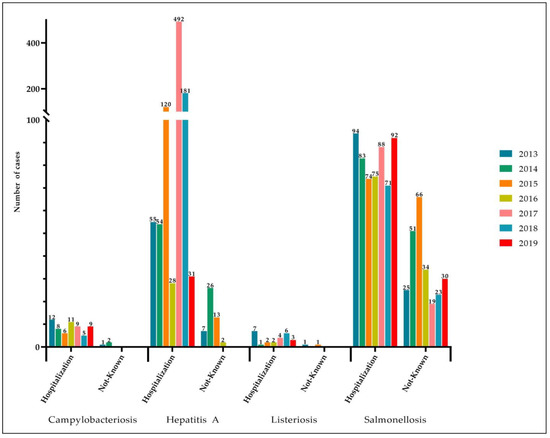





Considering the ability of Salmonella to survive on abiotic surfaces, this bacterium can represent a potential danger for consumers by contaminating food products. There is limited comprehension of the effect of surfaces, times, and temperatures used in food industries, as well as the influence of low-nutrient conditions, such as food contact surfaces that may have organic residue on adhesion and biofilm formation of Salmonella. It is crucial to describe bacterial adherence, the capacity to form biofilms, and the sanitizer resistance of Salmonella to design efficient control measures and hygiene procedures. Some strategies could directly inhibit virulence factors, such as adhesion, EPS secretion, flagella inhibition, and protein synthesis involved in bacterial metabolism or quorum sensing.
Salmonella biofilm formation continues with the irreversible adhesion caused by the secretion of EPS composed of polysaccharides, proteins, and DNA, which form the matrix biofilm and increase the cell-surface and cell-cell interactions [4]. The EPS matrix accounts for 90% of the biomass, while microorganism cells contribute the rest, 10%, emphasizing the significance of the EPS matrix. Salmonella biofilm’s main components consist of polysaccharides, cellulose, colanic acid, anionic O-antigen capsule, proteins such as the amyloid fibers called curli, flagella, surface protein components, and fatty acids [9]. The amount of each component within Salmonella biofilms is still unknown, representing an excellent area for further studies. It is essential to point out that the exact composition of the biofilms cannot be generalized for all cases; it would be interesting to know how the EPS components of the biofilm vary among serotypes or by modifying environmental factors and how this influences disinfection processes. For example, Kim et al. [45] concluded that the optimal condition for total cell mass and EPS synthesis after 9 days of Salmonella Typhimurium biofilm maturation was at 15 °C for the rdar (red, dry, and rough) and bdar (brown, dry, and rough) strains compared to 25 and 37 °C. It is necessary to establish the most specific contribution of each type of EPS in biofilm formation, and how this and the environmental factors influence the mechanisms of resistance to and survival of disinfectant processes, because only the contribution of the entire matrix is known.During biofilm maturation, the EPS matrix creates a three-dimensional network essential to biofilm lifestyle and virulence development; this network protects bacterial cells from environmental stresses, such as antimicrobials and immune system cells. The biofilm biomass provides a hydrated viscous environment that protects cells from various stressors, including desiccation, disinfectants, antibiotics, temperature, and oxygen content [46]. It may also prevent the loss of enzymes, nutrients, and molecules that could favor the microenvironment for bacteria within the biofilm [47]. However, the lack of knowledge detected on this point is regarding biofilm characteristics to consider its maturity level; these characteristics may involve measuring the number of adhered cells, EPS content, and variations among serotypes at different times. Salmonella’s lifetime is finished when cells leave, disperse from the biofilm, and revert to planktonic mode [48]. The dispersion of bacteria in biofilms should also be explored to a greater extent, because once they are released from the biofilm, they can colonize new sites and persist in the medium, making disinfection processes more difficult. Still, limited information is available on the time at which biofilm dispersal begins, as well as what environmental factors promote it.Structural EPS is the largest and most relevant group of substances that interferes in bacterial disinfection tasks. They primarily consist of neutral polysaccharides and protein parts that aid construction and surface colonization [49]. These EPS contribute to the formation processes, highlighting cellulose as one of the main components, followed by other components, such as curli, cholic acid, and protein O antigen material [9]. For all the stated above, biofilms are extremely difficult to remove from surfaces in the food industry. A lack of knowledge is detected around the potential variations in biofilm development, composition, and resistance among Salmonella strains with different morphotypes. In this context, it will also be interesting to determine the ideal conditions for biofilm formation and development. 5. Inhibition of Extracellular Polymeric Substances to Reduce Biofilm FormationThe traditional process to eradicate bacterial biofilms on food contact surfaces is cleaning and disinfection with conventional chemicals, such as chlorine, peracetic acid, and sodium hypochlorite [72]. However, current sanitation methods in the food industry have some well-known disadvantages, such as toxic residues of disinfection agents, corroded food contact surfaces, or the increasing resistance of these chemicals in microorganisms transmitted by food [73].The EPS matrix facilitates surface adhesion and colonization; therefore, the inhibition or disruption of EPS could be viewed as a target to prevent or eliminate biofilms. A promising concept to fight against biofilm infection is agents that could attenuate the production of those structural components inside the EPS matrix. Powell et al. [11] postulated that Pseudomonas aeruginosa preformed biofilms treated with oligosaccharides caused a significant decrease in the fluorescence intensity of ConA and TOTO-1 staining, indicating a biofilm disruption through the reduction in polysaccharides and extracellular DNA components of the matrix biofilm. Although the composition of Pseudomonas biofilms is different compared to Salmonella biofilms, the paradigm of targeting EPS and disrupting biofilms in Salmonella needs to be explored. Extracellular DNA (eDNA) has recently been discovered to be a novel element of Salmonella biofilms. eDNA was found and described in S. enterica ser. Typhi ST6 and 2-day-old S. Typhimurium SR-11 biofilms. According to reports, the eDNA acts as a biofouling agent. It is important to emphasize that the role of eDNA as an informative EPS component is not entirely established. More research must be done to fully comprehend how horizontal transmission could affect the development of biofilms’ development. As an alternative, the eDNA could help provide structural functions [74].It has been reported that curli-deficient S. enterica strains isolated from produce, meat, or clinical sources are the least effective in biofilm formation [75]. Moreover, curli is involved in Salmonella invasion, colonization, and persistent infections. The csgA gene’s deletion in S. enterica serovar Pullorum strain S6702 decreased curli production and biofilm formation, and reduced adhesion and invasion to HeLa cells’ intracellular proliferation in HD11 macrophages [27]. González et al. [76] reported that curli fimbriae are highly induced in a simulated human gallstone environment by developing biofilms on cholesterol-coated surfaces when bile was present. In a similar context, Adcox et al. [77] suggested a more significant contribution of curli in biofilm formation and gallbladder colonization than other extracellular matrix components.In contrast, curli is also crucial for attachment to plant surfaces. For example, curli was essential for transferring or surviving S. Typhimurium in parsley plants from contaminated irrigation water [78]. Brankatschk et al. [79] examined the whole transcriptome of Salmonella Weltevreden using RNA-seq and revealed that genes involved in the curli assembly were upregulated in the presence of alfalfa sprouts, suggesting an essential factor in colonization.Understanding curli fibers’ complex synthesis mechanism and their components allow the design of inhibition strategies against biofilm-related infections. For example, Yan et al. [80] used the dual-pore architecture of the CsgF-CsgG complex to create a peptide that physically reduces the size of the CsgF pore-inhibiting curli secretion and, thereby, curli amyloid fiber production in E. coli. Similar approaches have focused on preventing CsgA fiber polymerization. These studies demonstrated the activity of these treatments; however, there are still unknowns to be resolved and deepened. For example, some of these studies are in vitro, and evaluating more food systems or food contact surfaces would help us better understand biofilms’ behavior.Several studies have established that cellulose is involved in Salmonella biofilm formation and is essential for boosting resistance, which helps bacteria survive [45]. Yaron et al. [81] reported that cellulose of S. enterica is an adherence factor in plant materials. Bhowmick et al. [82] reported that deletion of gcpA in Salmonella enterica serovar Weltevreden caused the inability to produce cellulose. Consequently, the bacteria could not bind calcofluor to grow rdar colony on Congo Red-agar plates and develop biofilms on polystyrene surfaces.El Hag et al. [27] reported that deleting csgA and bcsA (deficient for curli and cellulose production) reduced biofilm formation in Salmonella Pullorum strain S60702 in glass tubes. However, ΔcsgA strain bacteria lowered adhesion and invasion into HeLa cells, but ΔbcsA did not. As cellulose is a crucial component of the biofilm matrix, inhibiting its production can be considered a control point for S. enterica biofilms. It is necessary to generate more knowledge regarding the terpenes’ effect on the genetic expression that encodes cellulose synthase. Similarly, post-translational studies are needed to understand the inhibition mechanisms of this enzymatic system; however, crystallizations of these proteins and structural characterization are required. Therefore, this enzyme could be a control point to inhibit cellulose synthesis and biofilm formation.As reviewed in this section, curli and cellulose are essential for Salmonella biofilms on different surfaces. Moreover, curli and cellulose-dependent biofilm are linked with several infections, including gastroenteritis [83]. Hence, developing agents to combat curli- and cellulose-dependent biofilms is a critical and pressing challenge. Studies suggested that EPS synthesis could be a biofilm control point, as the decrease in its content produces weaker and less structured biofilms, making them less resistant to conventional disinfectants. The identified knowledge gaps open a large area to explore agents that inhibit EPS synthesis and control biofilms in the food industry. 6. Terpenes as Potential EPS Inhibitors in Salmonella BiofilmsA promising alternative to prevent bacterial biofilm formation is the application of natural antimicrobials [8]. Terpenes and terpenoids, such as thymol, carvacrol, eugenol, and menthol, contained in essential oils, exhibit pronounced activities against diverse microorganisms (Table 2). Several essential oils and their main compounds have exerted antibiofilm activity against Salmonella [84]. For example, clove essential oil at 1.2 mg/mL decreased to 1.8 log CFU/cm2 of attached S. Typhimurium on stainless steel [15]. Carvacrol and thymol, phenolic components in oregano and thyme essential oils, exhibited antibacterial and antibiofilm action against S. Typhimurium [16]. Young and mature biofilms produced by S. Enteritidis on stainless steel surfaces were destroyed by essential oils of Origanum vulgare (0.25%) and Rosmarinus officinalis (4%), demonstrating a time-dependent impact and multitarget action mechanism on the bacterial membrane [17]. Čabarkapa et al. [19] investigated the antimicrobial and antibiofilm activities of Origanum vulgare, Origanum heracleoticum, Thymol vulgaris, and Thymol serpylluma versus S. Enteritidis. The results demonstrated that the essential oils and their principal constituents (carvacrol and thymol) prevented biofilm development at sub-MICs levels and eradicated 48 h formed biofilms over time in a concentration-dependent approach. Cinnamaldehyde (2 mg/mL), the major compound of cinnamon bark essential oil (55–76%), reduced the initial biofilm population by 6 log CFU/cm2 of Salmonella isolated from a conventional swine farm environment [85].However, few investigations on Salmonella have explored terpenes’ mechanisms to inhibit biofilms’ formation and eradicate them. Some studies in Table 2 have shown that the impairment of EPS synthesis is one of the mechanisms of phenolic compounds and terpenes in other bacteria. For example, carvacrol at 1.33 mM reduced the biofilm formation of Pectobacterium carotovorum—a bacterium that causes soft rot in plant food, attributed to its effect on EPS synthesis—for 24 h [88]. This compound at 0.66 mM (sub-inhibitory concentration) reduced the polysaccharides content in P. carotovorum biofilms from 22.94 to 9.11 glucose equivalent/cm2. At the same time, protein content was reduced, but to a lesser extent, from 11.07 to 7.10 albumin equivalent/cm2 [88]. Similarly, carvacrol at sub-inhibitory doses (64 and 128 μg/mL) reduced the biofilm formation of Enterobacter cloacae [20]. This effect was correlated with a decrease in biofilm exopolysaccharide production, which was observed in lower biofilm thickness and extra polymeric matrix by confocal laser scanning microscopy and scanning electron microscopy studies. In addition, the transcriptional study demonstrated that carvacrol down-regulated some genes, including curli fimbria and colonic acid polysaccharides necessary for biofilm formation in E. cloacae [20].In the same field, pimento berry, clove, and bay essential oils, and their main frequent component eugenol (0.0053 mg/mL), decreased the enterohemorrhagic E. coli O157:H7, an important foodborne pathogen worldwide [21]. The same study indicated that the eugenol structure, including the C-4 alkyl or alkane chain on the benzene ring, the C-2 methoxy unit, and the C-1 hydroxyl unit, have an essential function in antibiofilm mechanisms. This research also used transcriptional analysis to find that eugenol down-regulated some genes, such as fimbria type 1, fimbria curli, and cellulose (csgABDFG), in E. coli. The reviewed evidence indicated that plant terpenes affected EPS-secreting proteins in Gram-negative bacteria similar to the systems in Salmonella; therefore, we could hypothesize a similar effect.Hakimi Alni et al. [22] found that Cuminum cyminum essential oil, whose main compound is the terpenes terpineol, carene, and pinene, inhibited the growth of S. Typhimurium at 2.62 μL/mL. This essential oil at sub-inhibitory concentrations also reduced biofilm formation, and it caused the release of cells from biofilms, compared to control bacteria clinging together tightly and encased in a thick matrix. Moreover, these authors reported that the sub-inhibitory concentration of C. cyminum oil down-regulated significant genes related to biofilm formation, such as cellulose synthesis (csgD and adrA) and Quorum sensing (sdiA and luxS) genes.The clove essential oil, whose main component is eugenol, affected L. monocytogenes biofilm formation and removed biofilms on vegetable surfaces [24]. The results showed that all tested concentrations (0.5, 1.0, and 2.0 mg/mL) decreased 35–88% of extracellular polysaccharides and 34–76% of protein content in Listeria biofilms. In addition, this essential oil regulated the expression amounts of genes agrA, agrD, agrC, and prfA, and it upregulated the expression intensity of the gene sigB, thus controlling the formation of biofilms.Similarly, Li et al. [25] determined the effect of eugenol on glucan synthesis and biofilm of Streptococcus sobrinus, a caries-related pathogen. The results showed that an artificial mouth model’s minimum biofilm inhibitory concentration of eugenol (8 mg/mL) was 0.625 mg/mL. The reduction of insoluble glucan synthesis was 63% and 46% for soluble glucans. It is important to highlight that glucans are crucial for the biofilm of caries bacteria. However, many details of how this terpene inhibits glucan synthesis are not mentioned. Eugenol could potentially inhibit some enzymes in synthesizing glucans, such as glucosyltransferase. In addition, several studies conducted in this approach use oral bacteria, thus it would be essential to carry out additional studies that could elucidate the mechanism of glycosyltransferases in food pathogens.In this sense, Ortega-Ramirez et al. [26] found that citral and geraniol terpenes present in the Cymbopogon citratus essential oil inhibited planktonic growth (1.0 and 3.0 mg/mL) and biofilm formation of E. coli (2.0 and 4.0 mg/mL) on stainless steel. The mode of action of biofilm inhibition was attributed to the decrease in glucans production; citral and geraniol inhibited glucosyltransferase activity, demonstrating IC50 of 8.5 and 6.5 µM, respectively. Molecular docking showed that the most likely interactions between the terpenes and the enzyme occurred inside the hydrophobic pocket under the activation loop and near the enzyme’s finger helix. Terpenes modified glycosyltransferase activity based on the kinetic constants obtained; this suggested a mechanism of non-competitive inhibition of glycosyltransferase by citral and geraniol [26]. The terpenes’ interaction inside the hydrophobic bag beneath the activation circuit and the finger helix may influence UDP-glucose binding and glucan synthesis [89]. These authors describe a possible mechanism employing computational analysis of molecular couplings; therefore, complementing future studies, such as circular dichroism, RMN, and X-ray diffraction methods, could discover the mechanisms of action.Carrying out a brief analysis of the physicochemical characteristics of some terpenes, such as eugenol, carvacrol, and thymol, which are primarily a function of their structure that has an aromatic ring, it is suggested that this hydrophobic nature gives advantages to it to diffuse through the bacterial membrane in order to reach the intercellular space [90]. In addition, this molecule contains a polar part due to its hydroxyl group; this could allow it to interact with amino acids from the active site and modify the enzymatic action of glucosyltransferase. Once this molecule is in the intercellular region, it could interact with the catalytic subunit BcsA, which contains polar amino acids, such as aspartic acid, glutamine, or arginine, in the conserved domain [62]. These interactions could be by hydrogen bonds of the OH group of eugenol, with oxygen from the side chain of polar amino acids. However, the evidence that has been found to this day does not seem sufficient to decide what type of inhibition could occur and at which site of the enzyme; therefore, studies with this molecule and glucosyltransferase could clarify the information already mentioned above.In contrast to the inhibition of the cellulose-synthesizing enzyme, most studies evaluating curli synthesis inhibition are associated with terpenes’ effect on the gene expression of curli-related proteins. However, it is still necessary to delve into the mechanism to achieve this reduction. Furthermore, it would be enriching to research the exploration of the protein structures related to the curli, such as CsgA, CsgB, and CsgC, to design agents that possibly affect the assembly of fimbria amyloid. In addition, further studies need to evaluate whether terpenes directly influence these proteins, and how this affects adhesion to different surfaces and biofilm formation. This review indicates the lack of studies that demonstrate the potential of terpenes on the specific inhibition of colanic acid; the studies carried out evaluate EPS in a general way, and few studied the effect on each component of the biofilm.
Comments (0)