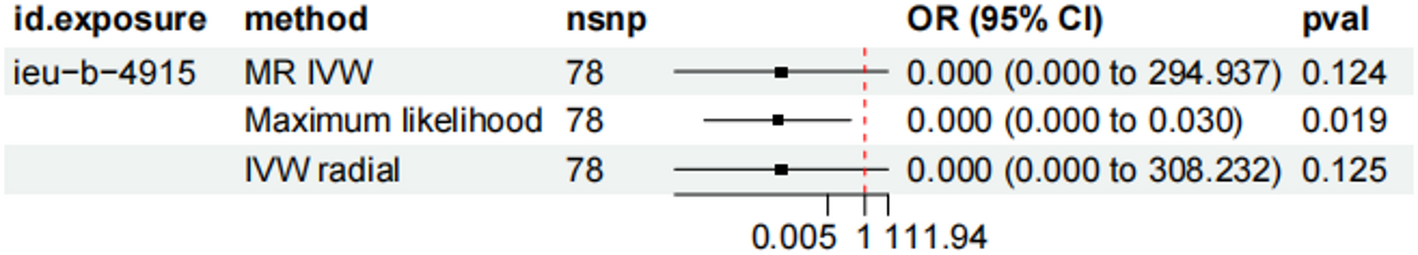
By integrating plasma proteomics with genome-wide association data, this study systematically identified plasma proteins associated with chronic lymphocytic leukemia through MR and Bayesian colocalization analyzes with the aim of uncovering potential drug targets. The discovery and evaluation of diverse therapeutic targets provides additional options for CLL treatment, including small molecule inhibitors, monoclonal antibodies, and other biologics. Nevertheless, individual patient responses may vary, highlighting the need to explore new strategies. Our MR analysis identified nine CLL-associated proteins, seven (CD177, SIGLEC5, MDGA2, LILRA1, HLA-C, MICA, PPIE) significantly increased CLL risk, while POGNT2 and CCL14 were associated with reduced risk. Colocalization analysis suggests that PPIE may be a promising therapeutic target. Further investigations using Phe-MR are planned to evaluate possible side effects and safety of targeted PPIE.
PPIE, an enzyme from the peptidyl prolyl isomerase (PPIase) family, was recognized as a CLL-related plasma protein in this study. The PPIase family plays a crucial role in various diseases. For example, PPIA, a prominent member called cyclophilin A (CyPA), facilitates the isomerization of proline residues in proteins. Previous studies using single-cell transcriptomics revealed high PPIA expression in primary refractory multiple myeloma patients and those unresponsive to KYDAR chemotherapy. CRISPR knockout of PPIA in the RPMI-8226 cell line showed that PPIA depletion or treatment with the CyPA inhibitor CsA increased sensitivity to the proteasome inhibitor CFZ, suggesting the role of PPIA in drug resistance and the potential for overcoming them [21]. B-cell malignancies such as multiple myeloma (MM) and CLL often colonize the bone marrow, where the microenvironment plays a critical role in their proliferation and protection [24,25,26]. Exogenous CyPA (eCyPA) secreted by bone marrow endothelial cells promotes proliferation and homing of MM cells by binding to the receptor CD147 [27].In addition to MM cells, eCyPA also promotes the migration of chronic lymphocytic leukemia (CLL) and lymphoplasmacytic lymphoma (LPL) cells, both of which express CD147 and tend to colonize the bone marrow.This suggests that PPIE, a member of the same family as PPIA, may play an important role in cellular signaling pathways. PPIE encodes an N-terminal RNA recognition motif (RRM) and a C-terminal isomerase domain [28]. It regulates chromatin modification, transcription and pre-mRNA splicing by directly interacting with the PHD3 domain of the histone reader MLL1 and associating with the XAB2 complex [29,30,31,32,33]. PPIE interacts with various splicing factors, including SF3B4, SF3A2, snRNPA’, B/B’ and SmD3, and may influence splicing patterns and thus protein production in B cells [34,35,36].
CLL is characterized by the pathological accumulation of mature B cells. Cellular stresses such as oxidative stress, DNA damage, and protein misfolding may contribute to disease progression by affecting apoptosis, cell cycle regulation, DNA repair mechanisms, and immune evasion. PPIE, as a molecular chaperone, is central to protein folding and catalyzes the isomerization of proline residues to ensure correct protein folding under stressful conditions, thereby preventing misfolding and protecting cells from damage [37]. The PPIase family also regulates the cell cycle and influences proliferation and aging, with CyPA being involved in the aging of hematopoietic stem cells. These potential functions and roles suggest that abnormal expression of PPIE plays an important role in the pathogenesis of CLL. However, further experimental validation is required to evaluate the potential of PPIE as a therapeutic target [38].
MICA, also known as MHC Class I Polypeptide-Related Sequence A, is a stress protein that is rarely expressed in normal cells, except in the gastrointestinal epithelium, endothelial cells and fibroblasts. High MICA expression is detected on various tumor cells, including non-small cell lung cancer, colon cancer, breast cancer and leukemia, which is consistent with our MR analysis. Tumor cells can evade immune surveillance by degrading MICA, leading to its internalization by NK cells via NKG2D, thereby silencing NK cell responses [39]. Therefore, anti-MICA antibodies represent a potential therapeutic target for CLL. CD177, also known as NB1 or HNA-2 antigen, is a GPI-anchored surface protein typically expressed on neutrophils. CD177 mediates neutrophil adhesion and migration through interactions with PECAM-1 and β2 integrin. It is considered a valuable marker in myeloproliferative diseases [40]. Although the association between CD177 and CLL risk has not been extensively studied, our findings suggest a potentially positive correlation. Studies have shown that CD177-deficient Treg cells reduce tumor growth and TI-Treg frequency in mice [40]. Although the strict correction thresholds are not met, several proteins with low p values are notable in our study. The SIGLEC protein family has been shown to contribute to the establishment of an immunosuppressive microenvironment in tumor cells by inducing a pro-cancer phenotype in tumor-associated macrophages and inhibiting immune cell activation [41]. Similar to CD177, there is limited research on the association of POGNT2 and CCL14 with CLL risk. Our MR results indicate a negative correlation between POGNT2, CCL14 and CLL risk, which warrants further investigation to understand the role of these three proteins in the development of CLL.
However, this study has limitations..Gene expression is highly complex and is influenced by various factors, including environmental conditions. The proteomic analysis in this study is limited to pQTL data from individuals of European ancestry, potentially introducing a bias for non-European populations. Furthermore, our main data source, plasma proteomics from the FinnGen cohort, is based on hypothesis-based genomic methods and does not include other relevant tissue systems, leading to selection bias. Unique pQTLs may have been missed, and unknown rare variants and complex counteractions may play an important role in genetics, warranting investigation in larger future studies.Regarding the assessment of druggability, it should be emphasized that this study identifies potential drug targets primarily through Mendelian randomization and colocalization evidence. There are currently no reports of drugs targeting this candidate for the treatment of CLL. Furthermore, the association of PPIE with various other clinical features raises concerns about possible off-target side effects, complicating its suitability as a drug target. Therefore, we conducted a protein interaction network analysis where alternative proteins within common signaling pathways may provide therapeutic options when the identified proteins are considered untreatable. This is because potential targets do not act in isolation but rather interact within a complex network of interconnected signaling pathways that contribute to disease pathology. In addition, some studies have shown that β2-microglobulin correlates with tumor burden, disease stage and prognosis in CLL, abnormalities in the secretion of plasma proteins such as IL-10 and TNF-α may be involved in the immune regulation of the tumor microenvironment and thereby tumor cell proliferation and affect survival. However, in the absence of confirmed tissue-specific roles in CLL, the use of plasma proteins as surrogates limits their explanatory power in directly implicating disease mechanisms. Therefore, although our method is based on careful hypotheses and considerations, it is crucial to integrate evidence from Mendelian randomization and colocalization, protein interactions with recently identified genes, as well as gene functionality and tissue-specific expression patterns to accurately determine disease-associated drug target genes. Furthermore, further clinical and microexperimental studies are essential to refine the evidence and confirm the potential of PPIE as a therapeutic target in CLL.




Comments (0)