
Remember me
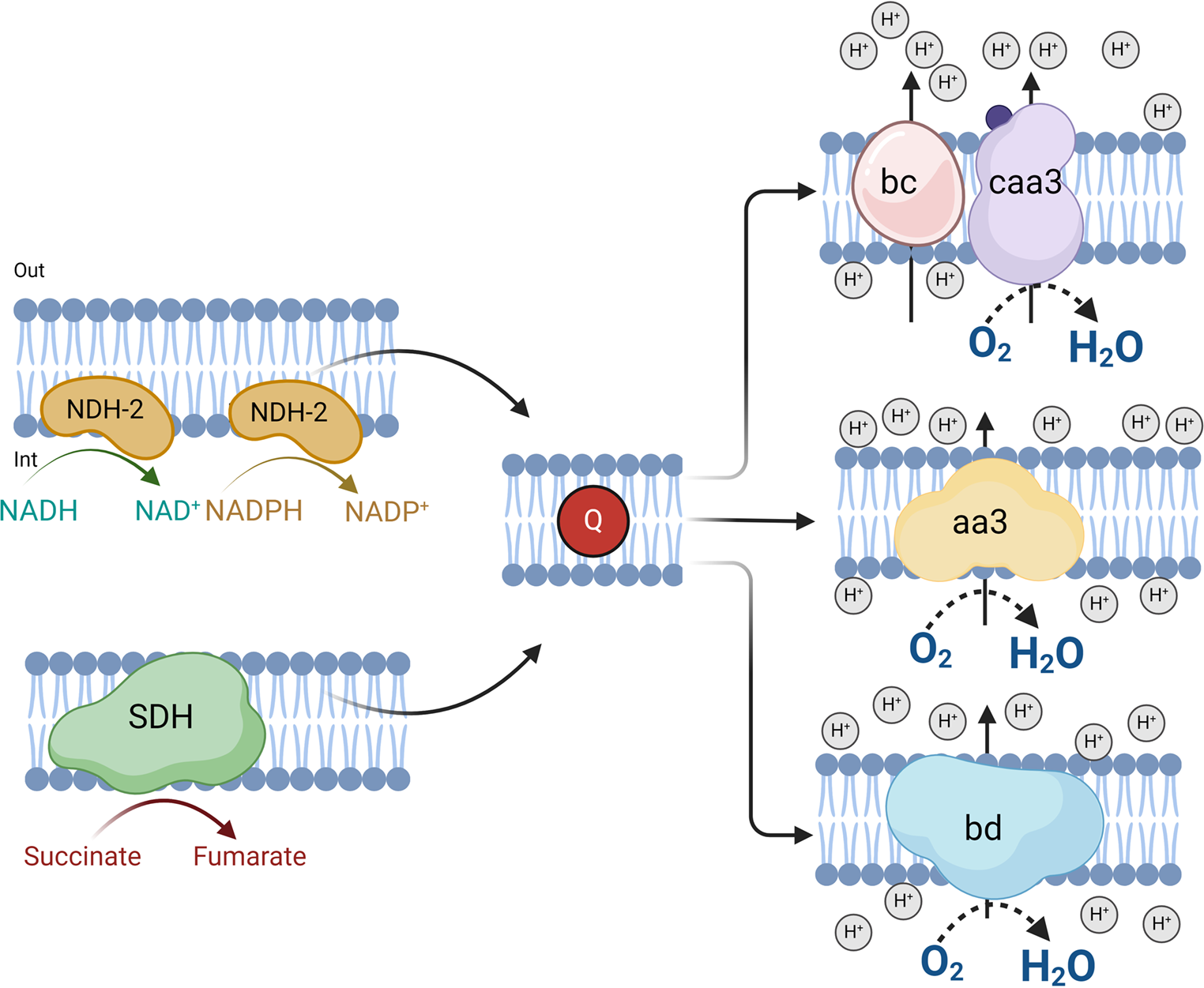
It has been reported that several bacteria, including B. licheniformis, can use cyanide as a nitrogen source (Mekuto et al. 2013; Rosario et al. 2023; Cáceda Quiroz et al. 2023). The degradation of cyanide associated with the production of ammonia was observed in batch cultures of B. licheniformis (Uribe-Ramírez et al. 2024). To further study the effect of cyanide on the growth of B. licheniformis, the bacterium was cultured in three different media: Nutritive Broth (Nut), an alkaline mineral minimum medium that contained ammonium (MM-NH4), and an alkaline mineral minimum medium with cyanide (MM-CN). Figure 2 shows the growth profiles of B. licheniformis at 25 ºC for 48 h in the three conditions studied. The results indicated that B. licheniformis could grow in the 3 culture media with different growth rates and biomass yields.
Fig. 2
Growth of Bacillus licheniformis in different culture media at 25 ºC and 140 rpm. a) Growth under the 3 culture conditions, b) plot of the growth curves in semi log scale
Cells grown in Nut medium reached a biomass concentration of 2.3 g L− 1 after 48 h incubation. The growth curves for MM-NH4 and MM-CN media began with approximately 1 g L− 1 of biomass to get enough material for the membrane isolation. B. licheniformis grew in MM-NH4 medium with a maximum specific growth rate of 0.061 h− 1 and reached a final biomass concentration of 2 g L− 1, implying a net biomass production of 1 g L− 1. In the case of the MM-CN medium, there was a lag phase of approximately 12 h, followed by an exponential increase in biomass with a maximum specific growth rate of 0.016 h− 1, and a final biomass concentration of 1.6 g L− 1 (net biomass production of 0.6 g L− 1) was reached at the end of the experiment. The highest maximum specific growth rate (0.167 h− 1) was obtained with the Nut medium. These results agree with those reported for Pseudomonas fluorescens with the same two nitrogen sources assayed in this work (NH4+ and CN−), obtaining better growth when ammonium was used as the nitrogen source (Harris and Knowles 1983). An explanation for the lower growth in the MM-CN condition compared to MM-NH4 is that the conversion of CN− into NH4+ is too slow, limiting the synthesis of biomolecules and the growth of the cells.
Oxygen consumption by B. licheniformis cellsOxygen consumption is an indicator of the metabolic status of the cell. A high respiratory rate indicates an abundance of respiratory enzymes and ATP synthase to support cell growth. Therefore, we studied the changes in oxygen consumption by B. licheniformis cultured in the three growth conditions tested (Fig. 3). The highest rate of oxygen consumption was observed in cells grown in Nut (546.23 nmol O2 min− 1 (mg dry weight)−1), followed by cells in MM-NH4 (102.64 nmol O2 min− 1 (mg dry weight)−1), and lastly by cells grown in MM-CN (49.74 nmol O2 min− 1 (mg dry weight)−1). The cell growth rate in the different media followed closely the respiratory rate. These results indicated a higher abundance of the respiratory elements in the Nut-cells.
Fig. 3
Oxygen consumption by B. licheniformis cells grown in the three different culture conditions assayed. The numbers in the figure indicate the oxygen consumption rates in nmol min− 1 (mg dry weight)−1
To determine the composition of the respiratory chain of B. licheniformis grown in the three culture conditions, we used specific inhibitors that interact with the respiratory enzymes in the plasma membrane. Flavone is well known to inhibit alternative NDH-2 dehydrogenases, and antimycin A and KCN interact with complex bc and cytochrome c oxidase, respectively. It is worth mentioning that in our experiments, the respiratory chain was fed with NADH, showing that the entry point of the electrons will depend on the activity of alternative NADH dehydrogenases.
Fig. 4
Effect of respiratory chain inhibitors on oxygen consumption of cells grown in Nut, MM-NH4, and MM-CN. Black bars represent oxygen consumption without an inhibitor, and empty bars represent oxygen consumption with an inhibitor. a) Flavone (960 µM), b) Antimycin A (28.8 µM), and c) KCN (1 mM)
Despite the high flavone concentration used in the assays, its inhibitory effect was small, at most 35% in the cells grown in the Nut medium. A smaller inhibition was observed for cells grown in MM-NH4 and MM-CN media (Fig. 4). Antimycin A did not affect the respiratory activity of the cells, regardless of the culture conditions (Fig. 4). Since antimycin A inhibits the bc complex, which is linked to the caa3 terminal oxidase, the results suggest the lack or minor amounts of these elements in the respiratory chain of B. licheniformis, at least under our culture conditions. For the cells grown in the Nut medium, cyanide inhibited respiration by around 90%. In contrast, for the cells cultivated in the MM-NH4 medium, the inhibitor did not significantly change their respiratory activity, and for the cells grown in the MM-CN medium, the addition of cyanide to the reaction medium increased oxygen consumption.
To further characterize the activating effect of cyanide on the respiratory activity of whole MM-CN-grown cells, we studied oxygen consumption at different cyanide concentrations. Figure 5 shows that the respiratory activity of MM-CN-grown cells depends on the cyanide concentration. It was found that the Michaelis-Menten model describes the relationship between oxygen consumption and the cyanide concentration, with a Vm of 8.0 nmol min− 1 (mg dry biomass)−1 and a Km for cyanide of 3.1 mM. Given that some reactions in the cyanide assimilation pathway involve oxygen consumption (Alvillo-Rivera et al. 2021), the simplest explanation is that the increase in oxygen consumption is due to reactions involved in cyanide metabolism. The observation that cyanide stimulated oxygen consumption in the cytosolic fraction of MM-CN-grown cells (Table 1) supports this interpretation.
Fig. 5
Oxygen consumption by cells grown in MM-CN at different potassium cyanide concentrations
Table 1 Oxygen consumption by cell-free extracts from MM-CN-grown cells at different cyanide concentrationsIt has been reported that an adequate supply of oxygen is important for the successful biodegradation of cyanide (Huertas et al. 2010). In this work, we demonstrate that the cyanide degradation metabolism of Bacillus licheniformis demands a high amount of oxygen.
Oxygen consumption in B. licheniformis membranesThe activating effect of cyanide on cellular respiration is expected to be absent in isolated membrane preparations, as the enzymes responsible for cyanide metabolism are localized in the cytosol rather than the membrane fraction. Therefore, membranes were isolated from cells grown in the three conditions, and their oxygen consumption was measured using NADH as a substrate. In addition, we tested the three mentioned inhibitors to look for the different respiratory enzymes.
Fig. 6
Oxygen consumption inhibition of cell membranes from Nut-, MM-NH4-, and MM-CN-grown cells. Black bars represent oxygen consumption without an inhibitor, and empty bars represent oxygen consumption with an inhibitor. a) Flavone (184 µM), b) Antimycin A (4.8 µM), and c) KCN (1 mM)
As shown in Fig. 6, oxygen consumption supported by NADH was highest in membranes from Nut-grown cells (290 nmol O2 min− 1 (mg protein)−1), followed by membranes from MM-NH4- (90 mol O2 min− 1 (mg protein)−1) and MM-CN (37 mol O2 min− 1 (mg protein)−1) grown cells. Similarly to the behavior of whole cells (Fig. 4), the inhibition by flavone was slight, regardless of the culture conditions. Oxygen consumption of membranes from Nut-grown cells was inhibited 30% by flavone and 40% in the other two conditions. Since the entrance of electrons to the respiratory chain depends on alternative NADH dehydrogenases, the partial inhibition of the oxygen uptake indicates that flavone might behave as a partial inhibitor, as described for the Ndi1 of S. cerevisiae (Velázquez and Pardo 2001). However, another explanation is that B. licheniformis NADH dehydrogenases are not inhibited by flavone. In agreement with the results obtained with whole cells, there was no inhibition by antimycin A, indicating that cytochrome bc and the caa3 terminal oxidase have a minor contribution to electron transport in this microorganism. Accordingly, the other two ubiquinol oxidases function as terminal oxidases.
Further information can be extracted from the conditions with cyanide. In agreement with the inhibition of oxygen consumption by whole cells, there was approximately a 60% inhibition by cyanide of membranes from Nut- and MM-NH4_grown cells. In contrast with the activating effect of cyanide in MM-CN-grown cells, in membranes from MM-CN-grown cells, there was 80% inhibition. Since the quinol oxidase aa3 is sensitive to high cyanide concentrations (Hill and Peterson 1998), the results suggest that 60 to 80% of the electron transport is due to the terminal oxidase aa3. In agreement with these data, it has been reported that cytochrome aa3 is mostly expressed in the respiratory chain of B. subtilis in the exponential phase (Winstedt et al. 1998). The remaining cyanide-insensitive (oxygen consumption in the presence of 1 mM cyanide) may originate from the bd-type terminal oxidases (Forte et al. 2017).
Several terminal oxidases have been described in Pseudomonas aeruginosa. It was reported that it has cytochrome bd, called Cio (cyanide-insensitive oxidase, encoded by the genes cioA and cioB), which is insensitive to cyanide (Luque-Almagro et al. 2011). Furthermore, Bacillus YN-2000 is a facultative alkaliphilic strain capable of growing in cyanide. The respiratory components of its plasma membrane were purified, and their oxygen consumption was measured and found to be 289 nmol O2 mg− 1 min− 1, which agrees with the oxygen consumption range of the Bacillus licheniformis cells used in this work. The oxygen consumption by membranes from Bacillus YN-2000 cells was 17.2 nmol O2 min− 1 (mg protein)−1 using NADH as a substrate and KCN (5 mM) as an inhibitor (Higashibata et al. 1998), comparable with what we report herein.
Specific activities of the B. licheniformis dehydrogenasesNext, we assayed some of the enzymes that feed the respiratory chain with electrons. Since the genome of B. licheniformis predicts several alternative NADH dehydrogenases and there is the possibility that one of these enzymes may use NADPH as substrate, we measured the dehydrogenase activity in isolated membranes with NADH and NADPH.
Fig. 7
Specific activity of membrane dehydrogenases obtained from Bacillus licheniformis grown under the three culture conditions with different substrates. a) NADH, b) NADPH, and c) succinate
As shown in Fig. 7, membranes obtained from Nut-grown cells showed NADH dehydrogenase activity, which was around 200 nmol min− 1 (mg protein)−1, twice the activity found with NADPH (Fig. 7b). These two activities were much lower in the membranes from MM-NH4- and MM-CN-grown cells, indicating a smaller amount of these enzymes in cells cultured in these two media. The activity with succinate (20 nmol min− 1 (mg protein)−1) was tenfold lower than that of the NADH dehydrogenase in the Nut-grown cells (Fig. 7c). Furthermore, the succinate dehydrogenase activity was only detected in membranes obtained from cells cultured in Nut and MM-NH4 media.
Activities of 200 and 100 nmol min− 1 (mg protein)−1 were reported for NADH dehydrogenases in Eikenella corrodens and Bacillus firmus, respectively. The NADH dehydrogenase activities in both microorganisms were comparable to that in B. licheniformis. However, succinate dehydrogenase activities reported in E. corrodens and B. firmus (235 and 420 nmol min− 1 (mg protein)−1, respectively) are 10 to 20 times the activity found in B. licheniformis (Hicks and Krulwich 1995; Jaramillo-Lanchero et al. 2021). So far, the activity of NADH dehydrogenase is comparable in the three species, but the succinate dehydrogenase activity in B. licheniformis is too small compared to that in E. corrodens and B. firmus. The growth conditions can explain these differences. In our experiments, cells were cultured in a medium containing 0.6% sodium acetate as a carbon source, while a rich medium containing 1.5% casein enzymatic hydrolysate, 1.5% sodium formate, and 0.1% of the following amino acids: cysteine, proline, methionine, tryptophan, and serine were used to grow E. corrodens (Jaramillo-Lanchero et al. 2021). B. firmus was grown in 50 mM malate as the carbon source (Hicks and Krulwich 1995).
Competition plotSince there was a possibility that the same enzyme was acting on NADH and NADPH as substrates, we relied on a simple kinetic technique, the competition plot (Fig. 8) to differentiate between one site binding the two substrates as opposed to separate sites. The competition plot involves plotting the total reaction rate against the parameter p, which represents the relative concentrations of the two substrates. If the two substrates react at the same site, the competition plot shows a horizontal straight line, indicating that the total rate is independent of p (Chevillard et al. 1993). However, if the reactions occur at separate sites, the plot exhibits a curve with a maximum. When NADH and NADPH were varied, a curve with a maximum was obtained (Fig. 8), indicating that the B. licheniformis membrane contains at least two enzymes, one specific for NADH and the other for NADPH.
Fig. 8
Competition plot for NADH and NADPH for NDH-2 of Bacillus licheniformis grown in Nut medium
Blue native gels of B. licheniformis respiratory enzymesUp to now, the respiratory activity of B. licheniformis depends on the activity of NADH, NADPH, and succinate dehydrogenases, but how these enzymes behave inside the membrane is still an open question. Are they forming complexes with other proteins? Are their molecular masses in agreement with the values calculated from the genome? In this regard, the association of the bc1 and caa3 complexes in B. subtilis and forming a bc1-caa3 supercomplex (García Montes De Oca et al. 2012), which might form respiratory strings in the membrane, has been reported (Picón Garrido et al. 2022). Additionally, a complex of the succinate dehydrogenase with the nitrate reductase has been proposed (Sousa et al. 2013). Therefore, the isolated membranes of B. licheniformis were incubated with digitonin to solubilize the proteins involved in the oxidative phosphorylation, then they were subjected to BN-PAGE, and the respiratory enzymes followed by specific in-gel activities. Approximately 150 µg of protein were loaded onto the gel for all conditions. Figure 9 shows the gel lanes with the activity of NADH dehydrogenase (Fig. 9a), NADPH dehydrogenase (Fig. 9b), succinate dehydrogenase (Fig. 9c), and cytochrome c oxidoreductase (Fig. 9d) for the three experimental conditions.
Fig. 9
Gel activities of digitonin-solubilized respiratory complexes from B. licheniformis grown in Nut, MM-NH4, MM-CN media. BN-PAGE 1D in a linear gradient of 4 to 10% acrylamide. a) NADH dehydrogenase, b) NADPH dehydrogenase, c) succinate dehydrogenase, and d) cytochrome c oxidase
Several bands of NADH dehydrogenase activity were detected in membranes from Nut- and MM-NH4-grown cells but not in the MM-CN-grown cells (Fig. 9), probably because of the low NADH-DH activity in cells grown in the presence of cyanide (Fig. 7). It seems that the activity in the gel appears when the specific activity of the NADH-DH is above certain limits (60–70 nmol min− 1 (mg protein)−1). Although we cannot discard the possibility that NADH-DH associates with other respiratory complexes or proteins, such as the bc-caa3 or the two quinol oxidases (aa3 and bd), the ladder banding pattern suggests the formation of homo-oligomers instead of the classic supercomplexes. Among the 5 NADH dehydrogenases found in the B. licheniformis genome and our proteomic study (Fig. 10; Table 2), only the enzyme with the accession number A0A1Y0YFU0 was associated with the high molecular weight bands containing NADH-DH activity (Bands 2 to 4 in Fig. 10; Table 2). The other NADH-DH were found near the bottom of the gel (Band 1 of Fig. 10; Table 2). Interestingly, the NADH-DH activity in B. subtilis appeared as a single band near the bottom of the gel, indicating that this enzyme could not associate with other proteins (García Montes De Oca et al. 2012; Picón Garrido et al. 2022) or form homo-oligomers. Gaining a better understanding of the NADH-DH aggregation mechanism in B. licheniformis will require the cloning, expressing, and purifying the enzyme.
The NADPH-DH activity in the gel was observed only in membranes obtained from Nut-grown cells (Fig. 9b). The protein’s molecular mass was around 173 kDa, suggesting a dimeric structure for the protein. In agreement with the competition plot (Fig. 8), NADH and NADPH dehydrogenase activities did not co-localize in the gel, further confirming the presence of two enzymes with different specificities for the coenzymes.
Membranes from Nut- and MM-NH4-grown cells, but not from MM-CN-grown cells, showed succinate dehydrogenase activity in the gel (Fig. 9c). The SDH migrated with a molecular mass of 375 kDa, much higher than the predicted from the genome (115 kDa). In agreement with the enzyme activity assays (Fig. 7), the in-gel activity for SDH was not detected in the MM-CN condition. It is worth mentioning that the bands containing the NADH, NADPH, and succinate dehydrogenase appear within 1 h incubation of the gel slices in the specific reaction mixtures. On the other hand, the cytochrome c oxidase band was observed only in the Nut condition after 48 h incubation of the gel slice with the reaction mixture, indicating a low activity of the caa3 complex.
2D native electrophoresisDigitonin is a mild detergent that extracts respiratory complexes and supercomplexes from the inner mitochondrial membrane while preserving the structural integrity of the supercomplexes (Cogliati et al. 2021). In contrast, the detergent DDM is a stronger agent that disassembles the supercomplexes’ components while maintaining the structure of the individual complexes (Lenaz et al. 2016).
Consequently, we solubilized the complexes using digitonin, performed a 1D BN-PAGE electrophoresis, incubated the gel slice with 0.02 or 0.1% DDM, and conducted a 2D BN-PAGE to separate the components of the respiratory complexes (Schagger et al. 1994). If the low molecular weight NADH dehydrogenase dissociates from the aggregates, its activity will appear near the bottom of the gel. Conversely, if the aggregates exhibit high stability, their migration will correspond to the molecular weight of the complex. If multiple stable complexes with NADH-DH activity exist, the activity will appear as a diagonal on the gel. Figure 10 shows that a significant portion of the NADH-DH activity dissociated from the high molecular weight complexes, likely corresponding to the dimeric form of the protein, and migrated to the bottom of the gel. Nonetheless, another fraction of the NADH-DH population remained as high molecular weight complexes and were distributed along the diagonal of the gel, suggesting strong interactions between the NADH-DH monomers.
Fig. 10
2D-BN-PAGE of digitonin-solubilized membranes from B. licheniformis grown in Nut medium previously resolved by 1D BN-PAGE. a) NADH dehydrogenase, b) NADPH dehydrogenase, and c) succinate dehydrogenase
For NADPH dehydrogenase, its migration indicates the high stability of the putative dimer. Interestingly, a less intense spot closer to the end of the gel also shows NADPH dehydrogenase activity, implying the existence of at least two enzymes capable of using NADPH as a substrate. Previous studies (García Montes De Oca et al. 2012) and our proteomic investigation (Band 5 of Fig. 11; Table 2) showed that the putative complex comprising the succinate dehydrogenase and the nitrate reductase was robust enough to withstand the high concentration of DDM.
In agreement with the studies in B. subtilis (Winstedt and Von Wachenfeldt 2000), our proteomic study (Table 2) revealed the presence of three terminal oxidases in B. licheniformis: the bc-caa3 and the two quinol oxidases, bd and aa3. Also, we did not find evidence for the presence of the bb’ oxidase (Azarkina et al. 1999). The experimental results showed that in B. licheniformis grown in the presence of acetate, the predominant terminal oxidases were bd and aa3, while the activity of bc-caa3 was negligible. This was consistent with the oxygen consumption results, where the bc-caa3 inhibitor antimycin A had no effect. Furthermore, when cells were cultured in the Nut medium, cyanide substantially reduced respiratory activity (90%), inhibiting the aa3 oxidase. In contrast, cyanide had a limited effect on cells grown in a minimal medium with acetate, indicating that the bd oxidase was the predominant form under this condition.
Fig. 11
1D-BN-PAGE of B. licheniformis membrane proteins. Bands B1 to B5 were analyzed by LC/MS-MS
Table 2 LC-MS/MS analysis of 1D BN-PAGE spots (1–5) identified in B. Licheniformis membranes from nut-grown cells
Comments (0)