
Remember me






The somatosensory system has functionally and anatomically distinct pathways conveying specific sensory modalities from the periphery to the brain.24,33,39,59 In a healthy state, somatosensory information (eg, touch, cold, and warm) drives approach and avoidance behaviors. However, pathological states can lead to abnormal sensory encoding of certain sensory modalities and, ultimately, to painful states.21,58,61 Patients with neuropathic pain display distinct sensory gain-of-function and/or loss-of-function profiles regardless of etiology, suggesting that there might be common mechanisms underlying the same sensory profile even from different pathologies.4,26,78 Clinical researchers have since turned to modality-specific assessment in peripheral painful neuropathies to reveal interactions between sensory phenotypes and the response to specific analgesics.2,7,16,17,25,42 Still, few links are known with specific neurobiological mechanisms, making targeted development of analgesics difficult.
By applying stimuli of different modalities (ie, mechanical, heat, and cold) in different transgenic lines, preclinical studies have made significant headway into describing circuitries linked to modality-specific nociception in normal and experimental models of pathological conditions.33,49 Approximately 70% of articles using animal models rely on reflexive assays, such as von Frey and Hargreaves tests.66 Although these methods can reliably detect whether certain nociceptive modalities are affected by a particular injury, they rely on entirely different physical quantities (ie, grams vs latency) that do not allow for direct comparison of modality-specific nociceptive behavior. Thus, one can hardly make conclusions about the relative importance of certain modalities in specific pathological conditions or the relative effect of certain analgesics on a specific modality.
Although these assays measure spinal reflexes, they do not consider the cognitive and emotional dimensions of pain. In the past decade, there has been a rise in the use of operant tests to measure voluntary pain behavior.32,66 Such assays use painful stimuli to alter reinforced behavior (ie, reward seeking or punishment avoidance) and assess the motivation to avoid certain stimuli.53,54,63 They are often limited to a single modality and rewards need to be carefully selected to avoid introducing sexually dimorphic preference/avoidance that could bias results.27,46,63 The conflict avoidance test was previously developed to obtain a detailed, stimulus–response assessment of pressure-evoked pain using light as an avoidance factor to cross pointed probes.23,28,70,71 Its use for modalities other than mechanical sensitivity has not been explored. A unified assay measuring mechanical, cold, and heat stimulus–response relationships on the same quantitative scale would offer new insights in mechanisms linked to certain phenotypes across pain models.
Our objective was to optimize and validate an operant and quantitative method based on the conflict avoidance paradigm to assess, using comparable parameters, modality-specific sensory behavior in mice. The dynamic range of stimulus–response relationships were determined for various outcome measures with mechanical, cold, and heat stimuli in naive animals. These measures were then used to estimate response profiles in nerve-injured animals and reveal differences in hypersensitivity across modalities and between the sexes.
2. Material and methods 2.1. AnimalsAll experimental procedures have been performed in accordance with guidelines from the Canadian Council on Animal Care and approved by the committee for animal protection of Université Laval (CPAUL; authorization number: 2018-027-3). Experiments were performed on adult (8-14 week old) male and female C57Bl/6J (JAX #000664; Jackson Laboratory or Crl : COBS.CD-1(ICR)BR colony (CD-1 mice, Charles River). Mice were housed in standard plastic cages in groups of 3 to 4 animals in ventilated racks under a 12-hours light–dark cycle starting at 7:00 am. Mice were allowed at least 1-week acclimatization to the facility after delivery before starting experiments. They were tested during light cycle, between 9:00 am and 6:00 pm. Testing was done in climate-controlled rooms (22-23°C, 30%-50% humidity). Behavioral assessment was conducted by Samuel Ferland (male experimenter).
2.2. Nerve injuryA mixture of isoflurane and oxygen was used during anesthesia (4% induction, 2% maintenance). In the cuff model, nerve injury was induced by placing a polyethylene cuff (0.38 mm ID, 1.09 mm OD, BD Intramedic PE20) around the sciatic nerve as previously described.5 In the spared nerve injury model (SNI), the tibial and common peroneal nerves were ligated using 6-0 silk sutures, then cut to leave the sural nerve intact.14 Baseline measurements for sensory tests were taken the week before surgery. Testing started 4 weeks after surgery.
2.3. Conflict avoidance assay 2.3.1. ApparatusThe apparatus (Coy MCS, Noldus, the Netherlands) consisted of a dark and a lit chamber linked by a corridor that we modified to deliver mechanical, cold, and heat stimuli in different sets of experiments (Fig. 1A). The dark and light chambers each measured 16.5 cm wide × 21.5 cm deep × 15.25 cm high. A mouse-adapted corridor (4.2-cm large, 4.2-cm high, and 39.5-cm long) linked the 2 chambers. The lighting in the light chamber was composed of a white 6.5 W LED bulb giving approximately 18,400 lux when on. Light intensities in the corridor and the dark chamber with the light on were of approximately 55.0 and 8.0 lux, respectively. Mice were acclimatized to the room for 60 minutes before tests, and all tests were done under red lighting in the room. A piece of white vinyl was put on the wall behind the corridor to improve the quality of video tracking. Testing order was randomized for each testing day by assigning a random number to each animal using the RAND() function in Microsoft Excel.
 Figure 1.:
Figure 1.: Validation of the effect of aversion to the initial compartment on training performances. (A) Diagram of the original and modified apparatus with measures of interest. (B) Diagram of the training protocol. (C and D) Percentage of the time spent in the initial compartment in the original and modified apparatus (n = 12 each), with and without light for (C) C57BL/6J mice (compartment F1,22 = 20.11, P = 0.0002, RM 2-way ANOVA) and (D) CD-1 mice (Compartment x light, F1,22 = 32.30, P <0.0001, RM 2-way ANOVA). Sex-separated values for CD1 are provided (D, right panel). Male mice are represented in black, and female mice in grey. (E) Difference between light off and on conditions for the modified compartment in both strains tested. (F and G) Comparison of escape and return latencies over the training procedure in the original (F), and modified (G) apparatus. Escape (H) and return (I) latencies of male and female mice trained in the original apparatus (Escape F1,18 = 3.739, P = 0.07; Return F1,18 = 10.73, P = 0.004, RM 2-way ANOVA). (J and K) Similar comparison for the modified apparatus (Escape F1,14 = 2.713, P = 0.1; Return F1,14 = 0.07963, P = 0.8, RM 2-way ANOVA). Data are shown as mean ± SEM. Differences between the groups are represented as asterisks (*), results from post hoc tests are represented as hashes (#), and results from the Mann–Whitney test are represented as a section sign (§). **P < 0.01, #P < 0.05, §§§P < 0.001, ####P < 0.0001, §§§P < 0.001, §§§§P < 0.0001, ns, not significant. ANOVA, analysis of variance; RM, repeated measures.
2.3.2. TrackingTrials were recorded using a USB webcam (Microsoft LifeCam Cinema). Experiments were planned, recorded, and run using Anymaze (Stoelting Co, Wood Dale, IL, version 6.34), but tracking had to be done post hoc using DeepLabCut (Mathis lab; version 2.2.0.3).44,52 DeepLabCut uses deep neural networks to estimate animal poses, thus providing better performances than traditional software on the videos with a nonuniform background. To train the model, we followed DeepLabCut recommendations and, specifically, we labeled the corners of the apparatus, the door, and the mouse center in 2904 images and used a ResNet50-based neural network for 5 training iterations (150-200 k iterations each, final test error = 2.77 pixels, train error = 2.20 pixels). Another model was trained to detect CD-1 with similar performances (test error = 3.29 pixels, train error = 3.07 pixels). These models were then used to extract positions from each video, and a modification of a preexisting code (https://github.com/FedeClaudi/DLCutils/blob/master/time_in_each_roi.py) was used to measure latency to escape from the light chamber, the latency to return from the dark chamber and the traversing speed (centimeters per second) for each trial.
2.3.3. Validation protocolTo validate the aversive nature of the light chamber, mice were allowed to explore the apparatus for 15 minutes with the lights off followed by another 15 minutes with the lights on. Time spent in the light chamber was measured with lights on and off and with or without a white background in the chamber.
2.3.4. Training protocolTo establish how training affected behavior, one mouse cohort underwent a 4-day training protocol without the presence of mechanical, cold, or heat stimuli. On the first day, they were acclimatized to the apparatus for 10 minutes (5 minutes lights off, 5 minutes lights on). In the following sessions (one session per day), they were placed in the dark chamber for 60 seconds, then in the light chamber for 60 seconds before the light was turned on and the doors were opened. Mice were allowed to escape to the dark chamber and return to the light chamber once. Trials were ended if animals stayed more than 60 seconds in any chamber without crossing. This was repeated for a total of 4 trials separated by at least 30 minutes per each session (Fig. 1B). Training was tested with and without white background in 2 separate cohorts of mice.
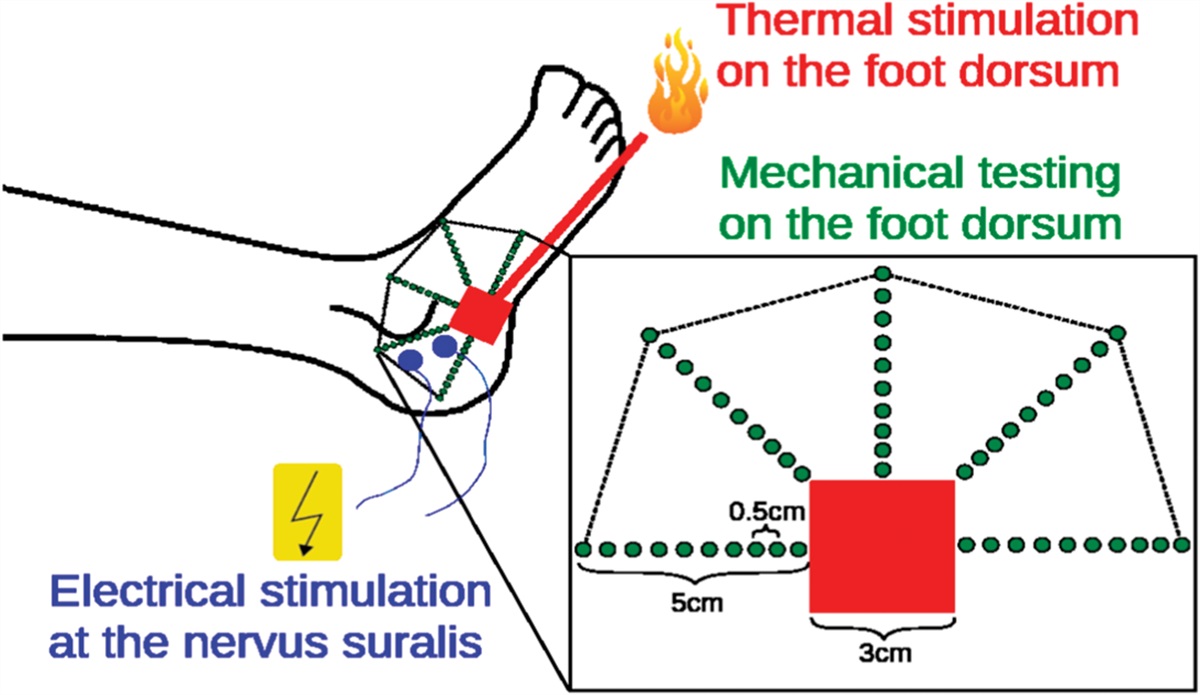
2.3.5. Testing protocolA different cohort of mice was tested with the presence of stimuli in the corridor. The testing protocol consisted of 5 days: acclimatization, training (1 session; see Results 3.1), and 3 testing days. Acclimatization and training were conducted as described above. On testing days, mice were presented with mechanical, cold, and heat stimuli. Two baseline trials (0 mm for mechanical, 30°C for thermal) were taken before pursuing with 3 stimuli of increasing intensity (raw baseline values are reported in Supplementary Fig. 2, available at https://links.lww.com/PAIN/B971). The cutoff point was increased to 120 seconds to allow for a wider range of behavior as previously used by others in mice.23,71 Mechanical stimuli consisted of a custom-made array of probes with adjustable height (0, 2, 3, 4 mm) separated by 5 mm. Thermal stimuli consisted of an aluminum plate placed on top of 2 hot–cold plates (Bioseb, France). Temperature was adjusted on the hot–cold plates so that the temperature of the aluminum plate reached wanted values (hot = 40°C, 45°C, 50°C; cold = 25°C, 20°C, 15°C). A piece of black electrical tape was affixed on the plate to measure temperature using an IR thermometer (Lasergrip 774, Etekcity). During test sessions, escape and return latencies, as well as traversing speed, were measured and expressed (in seconds and centimeters per second, respectively) as the difference from the group average measure of the 2 baselines taken on the same day. Because not all the tested animals escaped the light chamber or returned from the dark chamber within the cutoff time, the percentage of animals successfully crossing the corridor was also considered. Animals that crossed the midpoint of the corridor without reaching the dark chamber were excluded from the analysis of return successes.
2.4. Reflexive testsAnimals were acclimatized to the testing room for 60 minutes and to the apparatus for another 60 minutes before testing. Mechanical threshold was tested using the simplified up–down method6 with von Frey filaments #1 to #9.
2.5. StatisticsStatistical analysis was performed with Graphpad Prism 9 (Graphpad Software). An unpaired Mann–Whitney test was used to compare effect size after light exposure between C57BL/6J and CD-1 mice and results from the von Frey test. Three-way analyses of variances (ANOVAs) were used to compare the effect of sex, light, and compartment color on the time spent in the light compartment. Two-way ANOVAs were used to compare time spent in the light compartment, escape latency, return latency, and traversing speed between and within the groups. Repeated-measures (RM) 1-way ANOVAs followed by Bonferroni post hoc tests were used instead when only one variable was compared. Differences within and between the groups were analyzed using repeated-measures 2-way ANOVA followed by Bonferroni post hoc tests when they were appropriate. Within-group measures include light status, training session, or stimulus intensity. Between-group measures include the type of mechanical probes, sex, or surgery. Mixed-effects analysis was used instead when values were missing, such as when animals failed to escape the room and traverse the corridor. Geisser–Greenhouse correction was applied to correct for the violation of sphericity. The lowest P value between the main effect and the interaction was reported when results did not reach significance. Statistical differences in the number of successful vs unsuccessful escapes and returns from the light and dark rooms were assessed in a contingency table with Fisher exact test. P < 0.05 was considered statistically significant. The data are presented as mean ± SEM, with n representing the number of animals.
3. Results 3.1. Testing light aversiveness in the conflict avoidance apparatusThe conflict avoidance test uses a bright light to motivate animals to escape a chamber, traverse a corridor with noxious probes, and reach a dark chamber (Fig. 1A). The escape latency was previously shown to increase in the presence of mechanical probes in the corridor, and this increased further in nerve-injured rats.28 The aversive effectiveness of light in the initial chamber is therefore a key factor in driving the animals across the corridor, but light aversion was not investigated in the initial article. Therefore, we set out to investigate the aversiveness of the starting chamber, and how light affects performances after a 3-day training protocol (Fig. 1B). Surprisingly, we found that light was unable to induce strong aversive behavior in the original chamber, whose plexiglass walls were dark and transparent (Fig. 1C). Therefore, we decided to modify the light chamber by covering the walls with white vinyl stickers. C57BL/6J mice spent significantly less time overall in the modified chamber than the original one (Fig. 1C, P = 0.0002, n = 12 per group, RM 2-way ANOVA). Post hoc comparisons showed that light further decreased the time spent in the modified chamber (P = 0.04, Bonferroni post hoc test) but not the original one (P = 0.4). Conversely, the time in the dark compartment did not increase significantly after light exposure (Supplementary Fig. 1A, available at https://links.lww.com/PAIN/B971, P = 0.09). Because no sex-dependent effect on the time spent in the light chamber was observed (n = 6 per sex, P = 0.8, 3-way ANOVA), data from male and female mice have been combined. To test whether aversiveness was strain dependent, we also tested CD-1 mice in the original and modified apparatus (Fig. 1D). Unlike C57BL/6J, chamber configuration alone did not decrease the time spent in the initial chamber (Fig. 1D, P = 0.5, n = 12 per group, RM 2-way ANOVA), although light decreased it depending on the configuration used (P < 0.0001, eta-squared = 0.34). Post hoc comparisons showed that light significantly decreased the time spent in the compartment when it was delivered in the modified chamber (P < 0.0001). Comparison of the light aversion between C57BL/6J and CD-1 mice revealed that light had a bigger effect in CD-1 (Fig. 1E, P = 0.0004, Mann–Whitney test). Also, time in the dark compartment was significantly increased after light exposure (Supplementary Fig. 1B, available at https://links.lww.com/PAIN/B971, P = 0.0004) with a stronger effect for CD-1 than for C57BL/6J (Supplementary Fig. 1C, available at https://links.lww.com/PAIN/B971, P = 0.002, Mann–Whitney test). Sex had a small but significant effect on light aversion in the modified chamber (Fig. 1D, P = 0.03, eta-squared = 0.05, 3-way ANOVA), which can be explained by a higher preference of CD-1 female mice for the white chamber when the light is off. Still, both male and female mice decreased their time spent in the modified compartment when the light was turned on (males P = 0.01, females P < 0.0001, Bonferroni post hoc test).
Subsequently, we addressed training performances with both modified and original backgrounds for both escape and return latencies in C57BL/6J. Training was deemed successful when animals showed avoidance for the lit chamber (ie, shorter escape latency than return latency). The original chamber required more training sessions because escape latencies were shorter than return latencies only after the third training session (Fig. 1F, P = 0.0001, RM 2-way ANOVA with Bonferroni post hoc test). Conversely, using the modified chamber, escape latencies were shorter than return latencies after a single training session (Fig. 1G, P = 0.02). In these experiments, we also addressed differences because of the animal sex. Interestingly, sex differences were only observed with the original chamber, where returning times were longer in female mice than in male mice (Figs. 1H and I, P = 0.004, RM 2-way ANOVA). Instead, the modified chamber produced similar escape and return latencies in both sexes (Figs. 1J and K, escape P = 0.1, return P = 0.8). These data indicate that using a modified chamber with white walls provides stronger aversion, quicker training, and responses that are not affected by sex. Therefore, the modified chamber with a single training session was chosen for subsequent tests.
3.2. Testing mechanical sensitivity in mice with the conflict avoidance apparatusMice were tested for mechanical, cold, and heat sensitivity over a 5-day protocol (Fig. 2A). Mechanical sensitivity was originally tested by others in rats by measuring the avoidance induced by mechanical probes in the corridor against the aversiveness induced by light. Briefly, increasing the probe height was shown to increase the latency of the animal to escape from the light chamber.28 However, we noticed that although the original interprobe distance (1 cm) was well suited for the rats' paws, it was not adequate for mice. Therefore, to develop an apparatus suitable for mice, we designed 2 custom-made arrays of probes separated by 0.5 cm, one with nails and one with map pins (Fig. 2B, nail diameter = 500 µm, pin diameter = 125 µm). Then, escape and return latencies and traversing speed were compared in naive mice of both sexes exposed to either nails or pins with different heights. To facilitate comparisons between stimuli intensities, data are expressed as the difference between each measurement and the corresponding average baseline. Raw baseline values are reported in Supplementary Fig. 2A-D (available at https://links.lww.com/PAIN/B971).
 Figure 2.:
Figure 2.: Aversive response to mechanical probes of different sizes in naive male and female mice. (A) Diagram of the testing protocol for all modalities. (B) Microscopic images of nail and pin tips (left and right panel, respectively; scale bar = 500 µm). (C) Escape latency of male mice exposed to nails (open circles) or pins (full circles) (stimulus × probe height F3,130 = 8.105, P < 0.0001, 2-way ANOVA). (D) Return latency of male mice exposed to nails or pins (probe height F2.242,79.23 = 6.662, P = 0.001; stimulus F1,45 = 5.307, P = 0.03, 2-way ANOVA). (E) Escape latency of female mice exposed to nails or pins (stimulus F1,48 = 3.735, P = 0.06; Probe Height F2.470, 112.0 = 10.89, P < 0.0001). (F) Return latency of female mice exposed to nails or pins (stimulus × probe height F3, 114 = 3.532, P = 0.02, 2-way ANOVA). Data are shown as mean ± SEM. Differences from baseline for pins are represented as hashes (#). Scale bar = 500um. ##P < 0.05, ###P < 0.001, ####P < 0.0001. ANOVA, analysis of variance.
In male mice, pins produced a larger aversive effect as compared with nails for both escape latencies (Fig. 2C, P < 0.0001, n = 16 for nails and n = 24-32 for pins, 2-way ANOVA) and return latencies (Fig. 2D, P = 0.03, 2-way ANOVA). Post hoc comparisons revealed a significantly longer escape latency for pins set at 3 mm (P < 0.0001, Bonferroni post hoc test) and 4 mm (P < 0.0001) when compared with baseline (0 mm), which indicates an intensity-dependent aversive effect (Fig. 2C). Conversely, return latencies for pins were only significantly longer than baseline at 4 mm (Fig. 2D, P = 0.003). No differences from baseline were found for either escape or return latencies when animals were exposed to nails (Figs. 2C and D).
Also, in female mice, a stronger aversive effect was observed with pins rather than nails, although it was not significant in escape latency (Fig. 2E, P = 0.06, n = 18 for nails and n = 24-32 for pins, 2-way ANOVA) but only for return latency (P = 0.02). Indeed, probe height increased escape latency for both stimuli (Fig. 2E, P < 0.0001, n = 18 for nails, n = 24-32 for pins, 2-way ANOVA). Conversely, pins had a significantly stronger effect than nails on return latency depending on probe height (Fig. 2F, P = 0.02). Post hoc comparisons revealed that escape latency was longer with pins set at 3 mm (P = 0.003, Bonferroni post hoc test) and 4 mm (P = 0.0002) when compared with baseline (0 mm). Similar results were also found for return latencies with pins set at 3 mm (P = 0.003) and 4 mm (P < 0.0001). As for male mice, post hoc comparisons failed to detect significant differences with nails when compared with baseline.
When comparing aversion with mechanical stimuli between the sexes, we observed that female mice exposed to nails displayed longer escape latencies than male mice (P = 0.03, n = 16 for male mice, n = 18 for female mice, RM 2-way ANOVA), whereas no effect was observed on return latencies (P = 0.1). Contrastingly, both sexes had similar escape and return latencies when exposed to pins (escape, P = 0.7, return P = 0.2, n = 24-32 for each, 2-way ANOVA).
In addition to escape and return latency, we also measured the traversing speed on the first crossing (from light to dark chamber) and the second crossing (from dark chamber to light chamber). No differences were found in traversing speed between either stimulus for male or female mice. However, increasing probe height significantly decreased traversing speed for male and female mice (Supplementary Fig. 3, available at https://links.lww.com/PAIN/B971, P < 0.0001 for first and second crosses, RM 2-way ANOVA).
Altogether, pins were found to be more aversive than nails in both sexes and capable to induce a steeper stimulus–response relationship. Moreover, female mice appear more sensitive to mechanical stimuli than male mice in the conflict avoidance apparatus.
3.3. Testing thermal sensitivity in mice with the conflict avoidance apparatusTo allow testing thermal sensitivity, the floor of the corridor was equipped with a single aluminum plate placed upon 2 hot–cold plates for temperature adjustments. We then tested responses of male and female mice to temperatures ranging from innocuous to noxious cold and heat.
Cold stimuli induced a progressive increase in both escape and return latencies along with the lowering of the temperatures (Figs. 3A and B, P < 0.0001 for both, n = 16 for male mice, n = 18 for female mice, RM 2-way ANOVA and 2-way ANOVA). In male mice, both escape and return latencies were significantly longer at 20°C (escape: P = 0.04, return P = 0.0001; Bonferroni post hoc test) and 15°C (escape P < 0.0001, return P = 0.03; Bonferroni post hoc test) compared with baseline temperature. In female mice, longer escape latencies were only found at 15°C (P = 0.02), whereas return latencies were significantly increased at both 20°C (P = 0.01) and 15°C (P = 0.0002). The differences in escaping and returning behavior between the sexes were statistically significant (Escape P < 0.0001, Return P = 0.04, RM 2-way ANOVA and 2-way ANOVA). No relevant correlations with temperature and/or sex were found for the traversing speed on the first and second crossing (Supplementary Fig. 4A and B, available at https://links.lww.com/PAIN/B971).
 Figure 3.:
Figure 3.: Aversive response to various temperatures in naive male and female mice. (A) Escape and (B) return latency of male (blue) and female (light blue) mice in response to cold temperatures (escape sex × temperature F3, 96 = 15.45, P < 0.0001; Return Sex × Temperature F3, 78 = 2.974, P = 0.04, RM 2-way ANOVA and 2-way ANOVA). (C) Escape and (D) return latency of males (red) and females (pink) in response to hot temperatures (Return Temperature F2.674, 75.76 = 18.50, P < 0.0001, RM 2-way ANOVA; Return Sex F1, 29 = 0.3699, P = 0.6, RM 2-way ANOVA). Data are shown as mean ± SEM. Differences from baseline are represented as hashes (#) for male mice and section signs (§) for female mice. *P < 0.05, ****P < 0.0001, #P < 0.05, ##P < 0.01, ###P < 0.001, ####P < 0.0001, §P < 0.05, §§P < 0.01, §§§P < 0.001, ns, not significant. ANOVA, analysis of variance, RM, repeated measures.
Heat stimuli significantly increased return latencies (Figs. 3C and D, P < 0.0001, n = 16 male mice and 18 female mice, RM 2-way ANOVA) but had no effect on escape latencies (P = 0.6, RM 2-way ANOVA). Pairwise comparisons indicated that return latencies were significantly longer than baseline at 45°C for female mice only (female mice P = 0.05, male mice P = 1, Bonferroni post hoc test) and 50°C for both sexes (females P = 0.0009, males P = 0.002). No effect of sex on return latency was found (P = 0.4). Heat also significantly reduced traversing speed on the first (P = 0.05, RM 2-way ANOVA) and second (P = 0.0006, 2-way ANOVA) crossing (Supplementary Fig. 4C and D, available at https://links.lww.com/PAIN/B971). None of the above measurements highlighted relevant sex differences in aversive behavior to thermal stimuli. A separate cohort of male mice was tested to determine the effect of even higher temperature on escape latency. Heat was only found to increase escape latency when set at 55°C (Supplementary Fig. 5, available at https://links.lww.com/PAIN/B971, P = 0.02, 1-way ANOVA with Bonferroni post hoc test).
Only return latency represented reliable parameters to measure heat sensitivity, although both escape and return latency could be used for cold. Although male mice appeared more sensitive to cold stimuli than female mice, no sex differences were observed for heat stimuli.
3.4. Sex-specific hypersensitivity profiles in nerve-injured animals detected by the conflict avoidance apparatusWe next addressed whether the improved conflict avoidance assay can detect changes in aversive behavior to different types of stimuli in 2 different animal models of nerve injury: a chronic injury model based on the sciatic nerve constriction by cuff (cuff model) and the SNI. The development of mechanical allodynia, a classical hallmark of neuropathic pain,12 was firstly assessed by the von Frey test in both male and female mice (Fig. 4). Male and female mice that underwent cuff surgery had decreased mechanical thresholds when compared with naive animals (Fig. 4, P < 0.0001, n = 16 for both sexes and both groups, Mann–Whitney test), and mechanical thresholds were even lower after SNI (Fig. 4, male mice P = 0.006, n = 16; female mice P < 0.0001 n = 12).
 Figure 4.:
Figure 4.: Validation of hypersensitivity following surgery using the von Frey test, a classical reflexive method. Paw withdrawal threshold of male and female naive, cuff, and SNI mice. **P < 0.01, ****P < 0.0001. PWT, paw withdrawal threshold; SNI, spared nerve injury.
We then proceeded to test them in our conflict avoidance apparatus. Data are reported as difference with the average baseline values to facilitate comparisons between the models; however, raw baseline data are shown in Supplementary Fig. 2E-H, available at https://links.lww.com/PAIN/B971. Both cuff and SNI male mice displayed significantly increased escape latencies in response to pins compared with naive animals (Fig. 5A, Escape P < 0.0001, n = 24-32 for naives, n = 12 for cuffs, n = 16 for SNIs, 2-way ANOVA). Pairwise comparisons of escape latencies revealed longer escape latencies in cuff animals when pins were set at 4 mm (P < 0.0001, Bonferroni post hoc test), and SNI animals when pins were set at 2 mm (P = 0.03), 3 mm (P = 0.0001), and 4 mm (P < 0.0001). In addition, SNI animals had longer escape latencies than cuffs at 2 mm (P = 0.0006) and 3 mm (P = 0.02). Return latencies were also significantly longer after nerve injury (Fig. 5B, Return P = 0.0008, n = 24-32 for naive, n = 12 for cuffs, n = 16 for SNIs, 2-way ANOVA), although the low number of animals that failed to reach the dark chamber precluded any pairwise comparison (Fig. 5C). Indeed, we observed that for some nerve-injured mice, mechanical stimuli were so aversive that they failed to either escape or return within the cutoff time. In these cases, mice either remained at the corridor entrance, explored the pins with their front paws and whiskers, or even went back again after venturing out shortly. By comparing the number of mice that left the light or dark chambers vs those that did not in a contingency table, we found that the number of failed escapes and returns was significantly greater in nerve-injured mice than naive mice when presented with 3-mm pins for SNI only (Figs. 5C and D, Escape P = 0.003, Return = 0.0007, Fisher exact test) and with 4-mm pins for both cuff and SNI mice (Figs. 5C and D, Cuff Escape P = 0.005, Return P = 0.02; SNI Escape P = 0.0005, Return P = 0.002). In addition, less SNI mice successfully returned from 2-mm pins (Fig. 5D, P = 0.0006).
 Figure 5.:
Figure 5.: Aversive response of male mice to mechanical probes following nerve injury. (A and B) Escape and return latency of nerve-injured male mice in comparison to naive male mice (Escape Injury × Probe height F6, 163 = 10.09, P < 0.0001, 2-way ANOVA; Return Injury × Probe height F6, 83 = 4.296, P = 0.0008, 2-way ANOVA). (C and D) Percentage of escape and return success of naive and nerve-injured male mice in the presence of mechanical probes. Data are shown as mean ± SEM. Results from post hoc tests comparing naive vs cuffs (#), naive vs SNI (§), and cuff vs SNI (†). Results from Fisher exact test are represented by asterisks (*). *P < 0.05, **P < 0.01, ***P < 0.001, ####P < 0.0001, §P < 0.05, §§§P < 0.001, §§§§P < 0.0001, †P < 0.05. ANOVA, analysis of variance; SNI, spared nerve injury.
Cold temperatures only mildly affected escape latencies in nerve-injured male mice (Fig. 6A, n = 16 for naives, n = 12 for cuffs, n = 15 for SNIs, P = 0.03, RM 2-way ANOVA). Pairwise comparison revealed that escape latencies increased at 20°C in SNI but not in cuffs (SNI P = 0.04, Cuff P = 0.3, Bonferonni post hoc test). No effect on return latencies was observed (Fig. 6B, P = 0.8, 2-way ANOVA). Similarly, a larger number of SNI mice failed to escape at 20°C (Fig. 6C, P = 0.02, Fisher exact test), although no differences were observed for return successes (Fig. 6D). No effect of nerve injury on heat responses were observed (Figs. 6E and F, Escape P = 0.9, Return P = 0.1, n = 16 for naives, n = 12 for cuffs, n = 15 for SNIs, RM 2-way ANOVA and 2-way ANOVA). Conversely, the number of SNI mice that returned at 50°C was significantly lower than the other groups (Figs. 6Gand H, P = 0.01, Fisher exact test). These data show that male mice develop injury-dependent hypersensitivity profiles, with prominent mechanical hypersensitivity in both models and some thermal hypersensitivity detectable in SNI only.
 Figure 6.:
Figure 6.: Aversive response of male mice to thermal stimuli following nerve injury. (A and B) Escape and return latency of naive and nerve-injured male mice in response to cold temperatures (Escape Temperature × Injury F6, 120 = 2.273, P = 0.04, RM 2-way ANOVA; Return Injury F2, 40 = 0.2559, P = 0.8, 2-way ANOVA). (C and D) Percentage of escape and return success of naive and nerve-injured male mice in response to cold temperatures. (E and F) Escape and return latency of naive and nerve-injured male mice in response to hot temperatures (Escape Injury F2,40 = 0.09868, P = 0.9, RM 2-way ANOVA; Return Injury F2,38 = 2.109, P = 0.1, 2-way ANOVA). (G and H) Percentage of escape and return success of naive and nerve-injured male mice in response to hot temperatures. Data are shown as mean ± SEM. Results from post hoc tests comparing naive vs SNI are represented as section signs (§). Results from Fisher exact test are represented by asterisks (*). *P < 0.05, §P < 0.05. ANOVA, analysis of variance; RM, repeated measures; SNI, spared nerve injury.
Nerve-injured female mice also showed longer escape latency when exposed to mechanical stimuli as compared with naive mice (Fig. 7A, P = 0.001, n = 24-32 for naives, n = 16 for cuffs, n = 12 for SNIs, 2-way ANOVA). Pairwise comparison revealed that escape latency was increased in SNI exposed to 3-mm (P = 0.005) and 4-mm pins (P = 0.004, Bonferonni post hoc test). This was only significant in cuffs at 4 mm (P = 0.04). Comparing escape latencies between cuff and SNI female mice revealed a significant difference between them at 3 mm (P = 0.02). An interaction was also found between injury and return latency (Fig. 7B, P = 0.05, 2-way ANOVA). Pairwise comparison indicated significant differences between SNI and naive at 4 mm (P = 0.01). In addition, SNI had longer return latencies than cuffs at 3 mm (P = 0.05). Moreover, pins did not discourage cuff female mice to leave the light or dark chamber even at 4 mm (Figs. 7C and D, Escape P = 0.2, Return P = 0.2), although it did discourage SNI female mice to return at 4 mm (Figs. 7C and DP = 0.006, Fisher exact test). Contrary to male mice, both cuff and SNI female mice showed a marked increase in escape latencies for cold stimuli (Fig. 8A, P < 0.0001, n = 18 for naives, n = 16 for cuffs, n = 12 for SNIs RM 2-way ANOVA). Pairwise comparison indicated longer escape latencies for both cuff and SNI at 15°C (P < 0.0001 for both, Bonferroni post hoc te
Comments (0)